Your browser does not fully support modern features. Please upgrade for a smoother experience.
Submitted Successfully!
 +1 credit
+1 credit
Thank you for your contribution! You can also upload a video entry or images related to this topic.
For video creation, please contact our Academic Video Service.
| Version | Summary | Created by | Modification | Content Size | Created at | Operation |
|---|---|---|---|---|---|---|
| 1 | Jose E. Mesonero | + 1771 word(s) | 1771 | 2021-08-03 04:30:22 | | | |
| 2 | Peter Tang | Meta information modification | 1771 | 2021-08-13 03:05:04 | | |
Video Upload Options
We provide professional Academic Video Service to translate complex research into visually appealing presentations. Would you like to try it?
Cite
If you have any further questions, please contact Encyclopedia Editorial Office.
Mesonero, J.E. Neurotransmitter Dysfunction in IBS. Encyclopedia. Available online: https://encyclopedia.pub/entry/13115 (accessed on 07 February 2026).
Mesonero JE. Neurotransmitter Dysfunction in IBS. Encyclopedia. Available at: https://encyclopedia.pub/entry/13115. Accessed February 07, 2026.
Mesonero, Jose E.. "Neurotransmitter Dysfunction in IBS" Encyclopedia, https://encyclopedia.pub/entry/13115 (accessed February 07, 2026).
Mesonero, J.E. (2021, August 12). Neurotransmitter Dysfunction in IBS. In Encyclopedia. https://encyclopedia.pub/entry/13115
Mesonero, Jose E.. "Neurotransmitter Dysfunction in IBS." Encyclopedia. Web. 12 August, 2021.
Copy Citation
Irritable bowel syndrome (IBS) is a functional gastrointestinal disorder whose aetiology is still unknown. Most hypotheses point out the gut-brain axis as a key factor for IBS. The axis is composed of different anatomic and functional structures intercommunicated through neurotransmitters.
IBS
microbiota
visceral hypersensitivity
colorectal motility
1. Introduction
Irritable bowel syndrome (IBS) is defined as a functional gastrointestinal disorder, whose main symptoms are recurrent abdominal pain, changes in the frequency or characteristics of stool and abdominal distension. As a functional gastrointestinal disorder, IBS does not have a morphologic, metabolic, or neurologic aetiology. It is diagnosed using Rome IV clinical parameters. IBS can be classified in 4 different subtypes according to patient’s bowel habit: IBS with predominant constipation (IBS-C), IBS with predominant diarrhoea (IBS-D) and mixed-IBS which alternates between diarrhoea and constipation (IBS-M). Another type of IBS is called unclassified (IBS-U) [1], where individuals who do not fall into the other intestinal pattern categories are included.
IBS is considered the most prevalent gastrointestinal disorder; its prevalence is estimated to be around 10% to 15% of the population in Europe and North America. Despite its high prevalence, the physiopathology of IBS is still unknown. There are many hypotheses about IBS aetiology: psychosocial disorders, microbiotic alterations, hypersensitivity to some food, intestinal motility disorders, changes in visceral pain perception, or neurotransmitter alterations, creating a complex disorder of the gut-brain axis [2]. This axis is composed of intestinal microbiota, the intestinal epithelial barrier, neurotransmitters, the central nervous system (CNS), enteric nervous system (ENS), autonomic nervous system, and the hypothalamic-pituitary-adrenal axis. Together, all these components communicate bidirectionally (mainly through neurotransmitters), so intestinal signals can influence brain functions and vice versa. In fact, IBS patients show differences in brain activation areas in response to rectal distension and pain compared with healthy controls; suggesting that IBS patients lack central activation of descending inhibitory pathways [3]. Recent studies have reported alterations in brain networks and networks of interacting systems in the gut in IBS patients, evidencing a potential role of neurotransmitters on IBS pathophysiology [4]. On the other hand, psychosocial factors such as stress, anxiety, or depression, where neurotransmitters can play a key role, are considered risk factors for IBS and may even contribute to an exacerbation of IBS symptoms [5].
In recent years, many studies have focused on the association between IBS and changes in gut microbiota [6]. Gut microbiota can modulate host production of different neurotransmitters, as well as produce some neurotransmitters themselves [7]. Gut microbiota could play a role in the aetiology of IBS as they influence intestinal motility, gastrointestinal physiology, neurotransmitter levels, and behaviour. Actually, germ-free rats display a delay in intestinal peristalsis and that can be reverted by colonization with Lactobacillus acidophilus or Bifidobacterium bifidum [8]. IBS patients show perturbed microbiota composition, although there is no common microbiotic signature among IBS patients [9]. An increase of Firmicutes, especially Clostridium and Ruminococcaceae with a decrease of Bacteroidetes, particularly Bifidobacteria can be obtained in several mucosal and faecal samples from IBS patients [10]. Moreover, preliminary data suggest correlations of regional brain structural differences with gut microbial taxa [4].
The pathophysiology of IBS is incompletely understood, but it is well established that alterations in the gut-brain axis, altered CNS processing, motility disturbances and visceral hypersensitivity contribute to IBS aetiology. Other, less relevant or less studied mechanisms involved in IBS include genetic associations, alterations in gastrointestinal microbiota, cultural factors, and disturbances in mucosal and immune function [11]. Alterations in the gut-brain axis and differences in brain function are major contributing factors to IBS aetiology; however, the implications of key neurotransmitters such as norepinephrine (NE), serotonin, glutamate, GABA, and acetylcholine (ACh) in IBS are still unknown. The aim of this review is to evaluate the current evidence about neurotransmitter dysfunction in IBS and explore its potential therapeutic treatment. The Rome IV criteria for the diagnosis of IBS consist of abdominal pain associated with an alteration in either stool form or frequency, occurring for at least 6 months. Neurotransmitter dysfunctions could contribute to IBS and some of its most prevalent symptoms used for its diagnosis, grouped into two main aspects, visceral hypersensitivity and altered motility (Figure 1), although they may also be involved in other symptoms such as diet-related digestive disturbances, psychosocial disturbances, anxiety, depression, fatigue, hypertension, dyslipidaemia, etc. Therefore, targeting those dysfunctions may open novel lines for IBS management, taking into account, that these symptoms may also be indirect effects mediated by other biological and psychological factors.
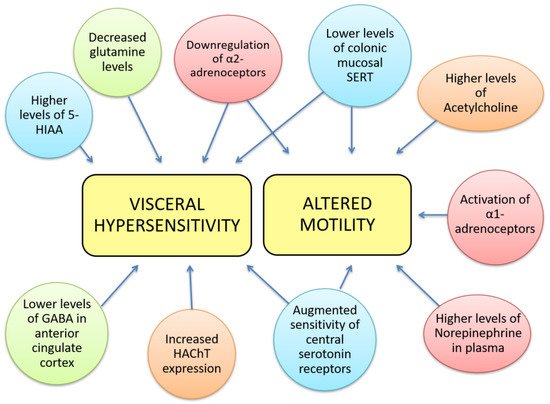
Figure 1. Neurotransmitter dysfunctions are related to some gastrointestinal IBS symptoms. Visceral hypersensitivity has been correlated to decreased glutamine levels, lower levels of GABA in the anterior cingulate cortex, higher levels of 5-hydroxy-indol acetic acid, increased expression of high affinity choline transporter, downregulation of α-2 adrenoceptors, augmented sensitivity of central serotonin receptors and lower levels of mucosal SERT. The latter 3 alterations can also be found in altered colorectal motility together with higher levels of NE in plasma, activation of α-1 adrenoceptors and higher levels of ACh. We notate neurotransmitter’s families with colours: red- norepinephrine; blue- 5-HT; green- GABA; orange-acetylcholinergic.
2. Norepinephrine
Noradrenaline (NE) is a key catecholamine with multiple physiological and homeostatic functions, key in the sympathetic nervous system. It is involved in excitation and the alert state during awake time, and in sensory signal detection. Secondarily, NE plays a role in behaviour, memory, attention, and learning. In fact, NE depletion in rats triggers distractibility and attentional deficits [12]. NE also has a leading role in spatial working, and memory functions, and its level is correlated with cognitive performance.
3. Serotonin
Serotonin was previously called enteramin by Erspamer and Asero due to its gastrointestinal functions; after discovering that it was identical to the vasoconstrictor substance known as such, it was renamed serotonin (5-HT, 5-hydroxytriptamin). Serotonin is synthesized from the amino acid tryptophan in enterochromaffin cells from the intestinal epithelium and serotonergic neurons. Ninety-five percent of serotonin production is from the gastrointestinal tract, meanwhile, 5% is from the nervous system. Once in the blood, serotonin can be stored in platelets, in which there are high levels of SERT (serotonin transporter) [13]. SERT uptakes 5-HT into cells, where it can be stored or degraded. SERT function is key to regulate 5-HT’s availability, and consequently 5-HT signalling.
Serotonin has multiple functions at the digestive level as a modulator of gastrointestinal secretion, peristalsis, or absorption; and also at a central level, controlling behaviour and critical neurological functions [14]. Experimental exogenous intake of serotonin results in multiple responses. This wide range of effects is due to the vast localization and diversity of 5-HT receptors [15]. Fourteen different 5-HT receptors have been identified and clustered in seven families based on their signalling pathways. Most of them are coupled to G proteins, and only the 5-HT3 receptor is a ligand-gated ion channel [16]. It is now known that some 5-HT receptors have specific functions, although many of them trigger diverse and antagonistic responses [17].
4. Glutamate
Glutamate is the main excitatory neurotransmitter in the CNS [18], and it has been described as having multiple roles as a nutrient, catalytic intermediate, or excitatory molecule [19]. Glutamate is an amino acid that can be introduced exogenously through the diet; however, exogenous glutamate crosses neither the intestinal barrier nor blood-brain barrier. Glutamate as a neurotransmitter is produced de novo in the brain from glucose [20]. After glutamate release from neurons, this amino acid is taken up by glia cells, and there, it is turned to glutamine by glutamine-synthetase for recycling to neurons. Glutamate reserves are refilled again when glutamine is engrossed by neurons. This means glutamine metabolism is the principal cycle for replacement of glutamate in neurons [21]. A high protein diet decreases glutamate and glutamine concentrations in plasma, although this phenomenon is still unexplained [22].
5. Gamma-Aminobutyric Acid
Gamma-aminobutyric acid (GABA) is an amino acid derivate of glutamate. Glutamic acid decarboxylase (GAD) enzyme is responsible for the conversion of glutamate to GABA by α-decarboxylation; afterwards, GAD interacts with the vesicular GABA transporter mediating the vesicular uptake of GABA [23]. Brain-derived neurotrophic factor (BDNF) increases GAD expression, regulating GABA homeostasis [24]. Ninety percent of the GABA synthesized is subsequently degraded by GABA-transaminase, which is present in neurons and glia cells. After its release from the nervous system, GABA transporter uptakes GABA from the synaptic cleft.
6. Acetylcholine
acetylcholine (ACh) is an excitatory neurotransmitter that is named after its chemical structure consisting of acetic acid and choline. Choline is present in dietary foods, and acetic acid derives from mitochondrial coenzyme acetyl-coA. The synthesis of ACh takes place in axon terminals and is catalysed by the enzyme choline-acetyl-transferase; then it is introduced in synaptic vesicles by the vesicular ACh transporter. After its release and binding to nicotinic or muscarinic receptors, ACh is degraded by acetylcholinesterase, mainly present in the synaptic cleft. Once hydrolysed, choline returns to presynaptic neurons by the action of a high-affinity choline transporter.
7. Other Neurotransmitters
The involvement of other neurotransmitters cannot be neglected. Several studies have pointed out the potential role of histamine and dopamine in IBS pathogenesis.
Histamine has been related to gastrointestinal inflammation and abdominal pain. The main histamine receptors, which take part in gastrointestinal processes, are H1 and H4, although H2 is related to the production of gastric acid [25]. In IBS patients, levels of urinary histamine have correlated to the severity of IBS symptoms, especially abdominal pain [26]. The administration of an H1-antagonist revealed different responses in IBS patients compared to healthy controls, demonstrating possible overstimulation of the histaminergic system in IBS patients [27]. H1 and H4 receptors could have a key role in the pathogenesis of colitis and postinflammatory visceral hypersensitivity, because their expression is increased in colon tissue of rats that have colitis. JNJ7777120, an H4-antagonist, seemed to ameliorate abdominal pain in that postinflammatory colitis model [28]. Novel interventions are being proposed that involve blocking H1 receptors, as ebastine has been found to improve IBS symptoms, including visceral hypersensitivity and abdominal pain [29], and ketotifen has been found to enhance health-related quality of life and increase the pain threshold in IBS patients [30]. Similarly, AST-12O, which adsorbs histamine from the intestinal lumen, could reduce pain and bloating in IBS-D and IBS-M patients [31].
On the other hand, several studies have investigated alterations in the dopaminergic system in IBS patients. In fact, IBS patients show lower dopamine levels in plasma [32] and urine [26] compared to healthy controls. Dopamine mediates colonic peristalsis, activating muscle contraction through D1 receptors and inhibiting it by D2 receptors [33], being related to motility dysfunction. However, the administration of dopamine or its agonists enhances IBS symptoms in patients with comorbid restless legs syndrome [34]. Nowadays, metformin is a widely used drug for the treatment of mellitus diabetes type II. Nevertheless, this drug has been studied for its antinociceptive effect through the activation of central D2 dopamine receptors in IBS patients [35]. Similarly, activation of those dopaminergic receptors by butyrate enemas decreases visceral allodynia and colonic hyperpermeability [36].
References
- Drossman, D.A.; Hasler, W.L. Rome IV-Functional GI Disorders: Disorders of Gut-Brain Interaction. Gastroenterology 2016, 150, 1257–1261.
- El-Salhy, M. Irritable bowel syndrome: Diagnosis and pathogenesis. World J. Gastroenterol. 2012, 18, 5151–5163.
- Tanaka, Y.; Kanazawa, M.; Kano, M.; Tashiro, M.; Fukudo, S. Relationship between sympathoadrenal and pituitary-adrenal response during colorectal distention in the presence of corticotropin- releasing hormone in patients with irritable bowel syndrome and healthy controls. PLoS ONE 2018, 13, e0199698.
- Mayer, A.E.; Labus, J.; Aziz, Q.; Tracey, I.; Kilpatrick, L.; Elsenbruch, S.; Schweinhardt, P.; Van Oudenhove, L.; Borsook, D. Role of brain imaging in disorders of brain-gut interaction: A Rome Working Team Report. Gut 2019, 68, 1701–1715.
- Surdea-Blaga, T.; Băban, A.; Dumitrascu, D.L. Psychosocial determinants of irritable bowel syndrome. World J. Gastroenterol. 2012, 18, 616–626.
- Wang, H.X.; Wang, Y.P. Gut Microbiota-brain Axis. Chin. Med. J. (Engl.) 2016, 129, 2373–2380.
- Labus, J.S.; Osadchiy, V.; Hsiao, E.Y.; Tap, J.; Derrien, M.; Gupta, A.; Tillisch, K.; Le Nevé, B.; Grinsvall, C.; Ljungberg, M.; et al. Evidence for an association of gut microbial Clostridia with brain functional connectivity and gastrointestinal sensorimotor function in patients with irritable bowel syndrome, based on tripartite network analysis. Microbiome 2019, 7, 45.
- Husebye, E.; Hellstrom, P.; Sundler, F.; Chen, J.; Midtvedt, T. Influence of microbial species on small intestinal myoelectric activity and transit in germ-free rats. Am. J. Physiol. Gastrointest. Liver Physiol. 2001, 280, 368–380.
- Simrén, M.; Barbara, G.; Flint, H.J.; Spiegel, B.M.; Spiller, R.C.; Vanner, S.; Verdu, E.F.; Whorwell, P.J.; Zoetendal, E.G.; Rome Foundation Committee. Intestinal microbiota in functional bowel disorders: A Rome foundation report. Gut 2013, 62, 159–176.
- Rajilić-Stojanović, M.; Jonkers, D.M.; Salonen, A.; Hanevik, K.; Raes, J.; Jalanka, J.; de Vos, W.M.; Manichanh, C.; Golic, N.; Enck, P.; et al. Intestinal Microbiota And Diet in IBS: Causes, Consequences, or Epiphenomena? Am. J. Gastroenterol. 2015, 110, 278–287.
- Ford, A.; Sperber, A.; Corsetti, M.; Camilleri, M. Irritable bowel syndrome. Lancet 2020, 396, 1675–1688.
- Borodovitsyna, O.; Flamini, M.; Chandler, D. Noradrenergic Modulation of Cognition in Health and Disease. Neural Plast. 2017, 2017, 6031478.
- Sebastián, J.J.; Sebastián, B. Serotonin and the two brains: Conductor of orchestra of intestinal physiology and mood role in irritable bowel syndrome. Med. Nat. 2018, 12, 11–17.
- Spohn, S.N.; Mawe, G.M. Non-conventional features of peripheral serotonin signaling Stephanie. Nat. Rev. Gastroenterol. Hepatol. 2017, 14, 412–420.
- Sharp, T.; Barnes, N.M. Central 5-HT receptors and their function; present and future. Neuropharmacology 2020, 177.
- Göthert, M. Serotonin discovery and stepwise disclosure of 5-HT receptor complexity over four decades. Part I. General background and discovery of serotonin as a basis for 5-HT receptor identification. Pharmacol. Rep. 2013, 65, 771–786.
- Green, A.R. Neuropharmacology of 5-hydroxytryptamine. Br. J. Pharmacol. 2006, 147 (Suppl. 1), S145–S152.
- Zhou, Y.; Danbolt, N.C. Glutamate as a neurotransmitter in the healthy brain. J. Neural. Transm. 2014, 121, 799–817.
- Fontana, A.C.K. Current approaches to enhance glutamate transporter function and expression. J. Neurochem. 2015, 134, 982–1007.
- Nakamura, E.; Uneyama, H.; Torii, K. Gastrointestinal nutrient chemosensing and the gut-brain axis: Significance of glutamate signaling for normal digestion. J. Gastroenterol. Hepatol. 2013, 28 (Suppl. 4), 2–8.
- Petroff, O.A.C. GABA and glutamate in the human brain. Neuroscientist 2002, 8, 562–573.
- Young, V.R.; Ajami, A.M. Glutamate: An amino acid of particular distinction. J. Nutr. 2000, 130, 892S–900S.
- Moloney, R.D.; O’Mahony, S.M.; Dinan, T.G.; Cryan, J. Stress-induced visceral pain: Toward animal models of irritable-bowel syndrome and associated comorbidities. Front. Psychiatry 2015, 6, 15.
- Meymandi, M.S.; Keyhanfar, F.; Sepehri, G.R.; Heravi, G.; Yazdanpanah, O. The Contribution of NMDA Receptors in Antinociceptive Effect of Pregabalin: Comparison of Two Models of Pain Assessment. Anesth. Pain Med. 2017, 7, e14602.
- Lieberman, P. The basics of histamine biology. Ann. Allergy Asthma Immunol. 2011, 106, S2–S5.
- Keshteli, A.H.; Madsen, K.L.; Mandal, R.; Boeckxstaens, G.E.; Bercik, P.; De Palma, G.; Reed, D.E.; Wishart, D.; Vanner, S.; Dieleman, L.A. Comparison of the metabolomic profiles of irritable bowel syndrome patients with ulcerative colitis patients and healthy controls: New insights into pathophysiology and potential biomarkers. Aliment. Pharmacol. Ther. 2019, 49, 723–732.
- Hattori, T.; Watanabe, S.; Kano, M.; Kanazawa, M.; Fukudo, S. Differential responding of autonomic function to histamine H 1 antagonism in irritable bowel syndrome. Neurogastroenterol. Motil. 2010, 22, 1284–1292.
- Deiteren, A.; De Man, J.G.; Ruyssers, N.E.; Moreels, T.G.; Pelckmans, P.A.; De Winter, B.Y. Histamine H4 and H1 receptors contribute to postin fl ammatory visceral hypersensitivity. Gut 2014, 63, 1873–1882.
- Wouters, M.M.; Balemans, D.; Van Wanrooy, S.; Dooley, J.; Cibert-Goton, V.; Alpizar, Y.A.; Valdez-Morales, E.E.; Nasser, Y.; Van Veldhoven, P.P.; Vanbrabant, W.; et al. Histamine Receptor H1-Mediated Sensitization of TRPV1 Mediates Visceral Hypersensitivity and Symptoms in Patients with Irritable Bowel Syndrome. Gastroenterology 2016, 150, 875–887.e9.
- Klooker, T.K.; Braak, B.; Koopman, K.E.; Welting, O.; Wouters, M.M.; van der Heide, S.; Schemann, M.; Bischoff, S.C.; van den Wijngaard, R.M.; Boeckxstaens, G.E. The mast cell stabiliser ketotifen decreases visceral hypersensitivity and improves intestinal symptoms in patients with irritable bowel syndrome. Gut 2010, 59, 1213–1221.
- Tack, J.F.; Miner, P.B.; Fischer, L.; Harris, M.S. Randomised clinical trial: The safety and efficacy of AST-120 in non-constipating irritable bowel syndrome—A double-blind, placebo-controlled study. Aliment. Pharmacol. Ther. 2011, 34, 868–877.
- Chojnacki, C.; Błońska, A.; Kaczka, A.; Chojnacki, J.; Stępień, A.; Gąsiorowska, A. Evaluation of serotonin and dopamine secretion and metabolism in patients with irritable bowel syndrome. Pol. Arch. Intern. Med. 2018, 128, 711–713.
- Zizzo, M.G.; Bellanca, A.; Amato, A.; Serio, R. Opposite effects of dopamine on the mechanical activity of circular and longitudinal muscle of human colon. Neurogastroenterol. Motil. 2020, 32, e13811.
- Prakash, S.; Prakash, A. Dopa responsive irritable bowel syndrome: Restless bowel syndrome or a gastrointestinal variant of restless legs syndrome? BMJ Case Rep. 2021, 14, e240686.
- Nozu, T.; Miyagishi, S.; Kumei, S.; Nozu, R.; Takakusaki, K.; Okumura, T. Metformin inhibits visceral allodynia and increased gut permeability induced by stress in rats. J. Gastroenterol. Hepatol. 2019, 34, 186–193.
- Nozu, T.; Miyagishi, S.; Nozu, R.; Takakusaki, K.; Okumura, T. Butyrate inhibits visceral allodynia and colonic hyperpermeability in rat models of irritable bowel syndrome. Sci. Rep. 2019, 9, 19603.
More
Information
Subjects:
Gastroenterology & Hepatology
Contributor
MDPI registered users' name will be linked to their SciProfiles pages. To register with us, please refer to https://encyclopedia.pub/register
:
View Times:
3.7K
Revisions:
2 times
(View History)
Update Date:
13 Aug 2021
Table of Contents



Notice
You are not a member of the advisory board for this topic. If you want to update advisory board member profile, please contact office@encyclopedia.pub.
OK
Confirm
Only members of the Encyclopedia advisory board for this topic are allowed to note entries. Would you like to become an advisory board member of the Encyclopedia?
Yes
No
${ textCharacter }/${ maxCharacter }
Submit
Cancel
Back
Comments
${ item }
|
${ item.createdUser.fullName }
${ item.createdAt }
${ item.vote }
${ item.reply }
Delete
${ reply.createdUser.fullName }
${ reply.createdAt }
${ reply.vote }
Delete
There is no reply to this comment~
${ item.replyTextCharacter }/${ item.replyMaxCharacter }
Submit
Cancel
More
No more~
There is no comment~
${ textCharacter }/${ maxCharacter }
Submit
Cancel
${ selectedItem.replyTextCharacter }/${ selectedItem.replyMaxCharacter }
Submit
Cancel
Confirm
Are you sure to Delete?
Yes
No