Your browser does not fully support modern features. Please upgrade for a smoother experience.
Submitted Successfully!
 +1 credit
+1 credit
Thank you for your contribution! You can also upload a video entry or images related to this topic.
For video creation, please contact our Academic Video Service.
| Version | Summary | Created by | Modification | Content Size | Created at | Operation |
|---|---|---|---|---|---|---|
| 1 | Przemysław Dariusz Gilun | + 1964 word(s) | 1964 | 2021-07-26 10:40:11 | | | |
| 2 | Amina Yu | Meta information modification | 1964 | 2021-08-05 09:35:03 | | |
Video Upload Options
We provide professional Academic Video Service to translate complex research into visually appealing presentations. Would you like to try it?
Cite
If you have any further questions, please contact Encyclopedia Editorial Office.
Gilun, P. Methylation in Period2 (PER2) Transcription. Encyclopedia. Available online: https://encyclopedia.pub/entry/12813 (accessed on 08 February 2026).
Gilun P. Methylation in Period2 (PER2) Transcription. Encyclopedia. Available at: https://encyclopedia.pub/entry/12813. Accessed February 08, 2026.
Gilun, Przemysław. "Methylation in Period2 (PER2) Transcription" Encyclopedia, https://encyclopedia.pub/entry/12813 (accessed February 08, 2026).
Gilun, P. (2021, August 05). Methylation in Period2 (PER2) Transcription. In Encyclopedia. https://encyclopedia.pub/entry/12813
Gilun, Przemysław. "Methylation in Period2 (PER2) Transcription." Encyclopedia. Web. 05 August, 2021.
Copy Citation
It has been proposed that carbon monoxide (CO) is a chemical light carrier that is transferred by the humoral pathway from the retina to the brain. Here, we aimed to study how deeply CO is involved in regulating the expression of Period2 gene (PER2), one of the genes maintaining the intrinsic biological clock.
gene expression
E-box
methylation
PER2
carbon monoxide
oscillation
1. Introduction
Many physiological processes are cyclic and depend directly or indirectly on the intrinsic biological clock. The main biological clock in mammals is located in suprachiasmatic nuclei (SCN) in the pre-optic part of the hypothalamus and is modulated endogenously by clock-related genes such as PER1, PER2, PER3, CRY1 and CRY2. These genes are regulated by a heterodimer of the specific transcription factors BMAL1/CLOCK that react with E-box enhancers in the promoter region of genes (PER1–3 and CRY1–2), creating a core loop. The PER and CRY proteins in turn heterodimerize and translocate to the nucleus to inhibit BMAL1/CLOCK activity. This is a cell-autonomous molecular oscillator containing a transcription-translational feedback loop (TTFL) that displays a roughly 24 h cycle. Circadian rhythm depends on external cues called zeitgebers, and the most important is light [1][2][3]. Sunlight falling on the retina starts the process of neural light transmission to the hypothalamus to SCN [4]. It has been reported that the retina can simultaneously produce carbon monoxide (CO) as a chemical light carrier and that this production depends on light intensity [5][6][7]. Further CO is transferred via the humoral pathway to the hypothalamus and modulates the biological clock as a „chemical light signal” independently from a neural light signal [8].
The Period2 gene is a member of the Period family (PER1–3). These genes cooperate with cryptochromes in TTFL in SCN to form the main biological clock. PER2 is also expressed in peripheral tissues, creating peripheral clocks that, in turn, depend on the main biological clock [9]. PER2 plays a more dominant role than PER1 in the circadian rhythm that affects the central and peripheral nervous systems [10][11]. Furthermore, Per2/PER2 is involved in many such processes in the nervous system, such as neurotransmitter regulation (GABA, dopamine, glutamate and serotonin) and prolactin production, as well as sleep and food anticipation. Some authors indicate that PER2 plays a role in neurodegenerative diseases, drug addiction and depression [11][12]. PER2 is regulated through the promoter sequences containing enhancer E-boxes [CANNTG] [13] and D-box (daytime expression enhancer) and also CRE (cAMP-response element) [14][15] in gene sequences downstream from ATG (start of translation).
Functional clock gene proteins such as BMAL1, CLOCK and NPAS2, as well as PER2, REV-ERB α and β belong to a group of proteins called basic helix-loop-helix PER-ARNT-SIM (bHLH-PAS), containing PAS A and PAS B domains that form functional protein dimers with different biological functions. These domains are able to bind heme groups with Fe(II)-valent iron, which acts as a cofactor and influences the function of these proteins. Binding heme groups to Fe(II) at the active center sensitizes these proteins to CO, which, by binding to heme, exerts such biological effects as inhibiting the dimerization of BMAL1:NPAS2 proteins [16][17][18].
The aim of this research was to study the regulation of PER2 expression over a 12 h period. We used pigs for the experiment, because they have a similar circadian rhythm as humans, in which daylight activity and housing with artificial light influence the biological clock. The pig brain is big enough for us to obtain an appropriate amount of samples from the hypothalamic preoptic area with SCN but without other nuclei; i.e., paraventricular (PVN in the dorsal part of the hypothalamus) or arcuate (ARC in the ventral part of the hypothalamus). We used CO as a chemical light signal because our recent research showed that the CO level in blood depends on time and light intensity and that it can modulate the biological clock in the hypothalamus [5][8]. Our experimental model assumed that only a chemical light signal or lack of it will be a cue for the organism to synchronize the main biological clock, which is why we used animals adapted to constant darkness.
We hypothesized that carbon monoxide delivered by the humoral pathway to the hypothalamus will change the PER2 promoter methylation in order to organize the mechanistic function of the main biological clock.
2. Role of Methylation in Period2 (PER2) Transcription in the Context of the Presence or Absence of Light Signals: Natural and Chemical—Studies on the Pig Model
2.1. PER2 Gene Expression
The PER2 mRNA expression in the hypothalamus was 2.5-fold higher (p < 0.0001) during the day in the normal group; this kind of pattern of daytime expression is called oscillation. In the control group, we did not observe statistically significant changes between day and night, and oscillation was damped. The PER2 expression trend reversed in the CO-treated group; during the night, it was significantly higher (p < 0.05) compared to expression during the day, and oscillation reversed at night (Figure 1).
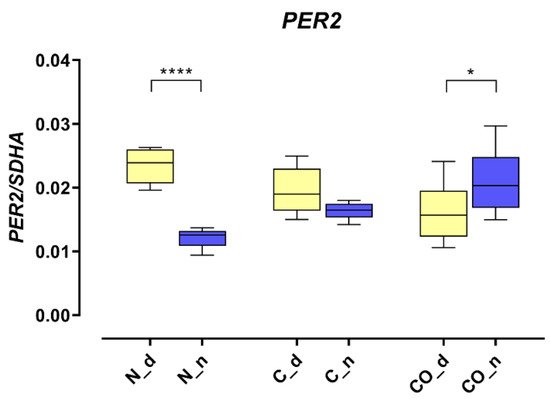
Figure 1. The PER2 mRNA expression in the preoptic part of the hypothalamus that contained suprachiasmatic nuclei in the normal (N), control (C) and CO-treated groups. The data were normalized by dividing the expression of the target gene by average expression of the housekeeping gene SDHA (succinate dehydrogenase). The bars are represented as mean ± SEM in arbitrary units. The PER2 mRNA expression obtained in the tissues of the hypothalamus during the day (yellow whisker boxes) and night (blue whisker boxes) were compared in particular studies groups. The asterisks indicate significant differences as follows: *—p < 0.05; ****—p < 0.0001.
2.2. Methylation Level
Four different promotor sequence regions were studied: −1–−79 bp, −1185–−1224 bp, −2000–−2054 bp and −2402–−2464 bp. The region closest to the ATG (−1–−79 bp) canonical E-box was hypomethylated. The DNA methylation was around 10%. In this region, the difference between night and day methylation levels was observed in the normal group at the 3rd (p < 0.01), 5th (p < 0.05) and 8th (p < 0.05) CpG positions. The CO-treated group showed a change in the methylation level at the 3rd CpG position. All the described hypothalamus samples exhibited a higher methylation level during the day. In the control group, the methylation level did not differ at any of the CpG positions (Figure 2). The highest methylation level (80–100%) was observed between −1185–−1224 bp. The methylation level was significantly higher at the 2nd CpG position in hypothalamic DNA samples collected at night in the normal (p < 0.001) and CO-treated groups (p < 0.01). The methylation level in the normal group at the 4th CpG position was also higher at night (p < 0.01) (Figure 3). The −2000–−2054 bp region methylation level was high at 60–80% and varied significantly between day and night only in the normal group at the 1st CpG position (p < 0.001) (Figure 4). The methylation level between −2402–−2464 bp in all the experimental groups was approximately 60%. There were no differences in methylation between day and night in this region (Figure 5).
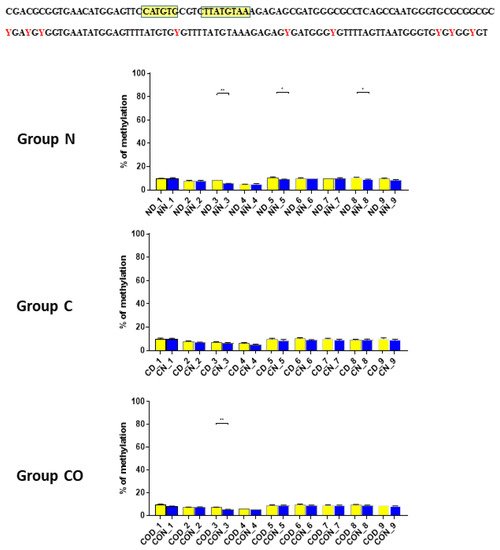
Figure 2. The difference between day (yellow bar) and night (blue bar) methylation levels in the hypothalamus in the normal (N), control (C) and CO-treated groups. The region closest to the ATG (−1–−79 bp) canonical E-box was hypomethylated. In this region, the difference between methylation was observed in the normal group at the 3rd, 5th and 8th CpG positions. The CO-treated group showed a significant change in the methylation level in the 3rd CpG position. The bars are represented as mean ± SEM. The asterisks indicate significant differences as follows: *—p < 0.05; **—p < 0.01. At the top of the figure is the examined sequence: original (top) with yellow highlighted sequences E-box and D-box and bisulfite converted with highlighted methylation position (bottom).
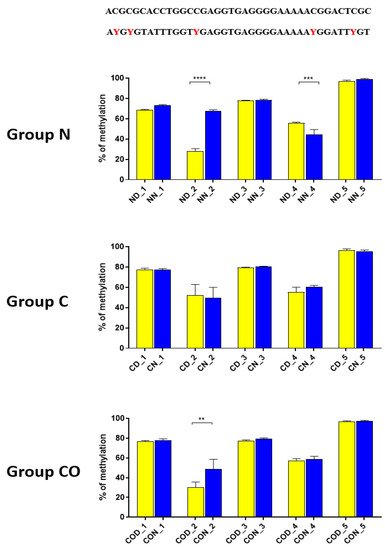
Figure 3. The difference between day (yellow bar) and night (blue bar) methylation levels in the hypothalamus in the normal (N), control (C) and CO-treated groups. The highest methylation level was observed in −1185–−1224 bp, where it was on 80–100%. The methylation level was significantly higher at the 2nd and 4th CpG position in hypothalamic DNA samples collected at night in the normal and in 2nd CpG position in the CO-treated group. The bars are represented as mean ± SEM. The asterisks indicate significant differences as follows: **—p < 0.01; ***—p < 0.001; ****—p < 0.0001. At the top of the figure is the examined sequence: original (top) and bisulfite converted with highlighted methylation position (bottom).
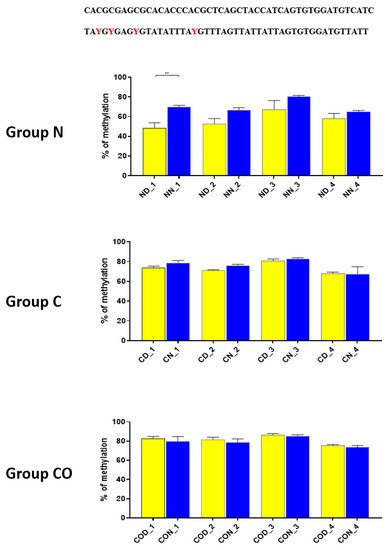
Figure 4. The difference between day (yellow bar) and night (blue bar) methylation levels in the hypothalamus in the normal (N), control (C) and CO-treated groups. The −2000—−2054 bp region methylation level was high at 60–80% and varied significantly between day and night only in the normal group in position 1 of CpG. The bars are represented as mean ± SEM. The asterisks indicate significant differences: **—p < 0.01. At the top of the figure is the examined sequence: original (top) and bisulfite converted with highlighted methylation position (bottom).
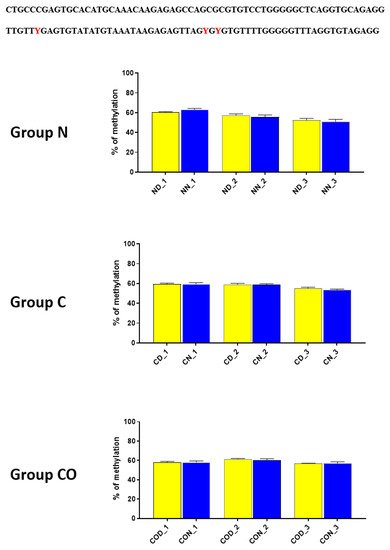
Figure 5. The difference between day (yellow bar) and night (blue bar) methylation levels in the normal (N), control (C) and CO-treated groups of the hypothalamus. The methylation level between −2402–−2464 bp in all the experimental groups was approximately 60%. There were no differences in methylation levels between day and night in this region. The bars are represented as mean ± SEM. At the top of the figure is the examined sequence: original (top) and bisulfite converted with highlighted methylation position (bottom).
We also analyzed the overall methylation level in each of the four regions (Figure 6) and observed that daily oscillation in region −1–−79 bp differed statistically (p < 0.05) between day and night in normal and CO-treated groups. In the control group, neither daily oscillation nor differences between groups were observed (Figure 6A). We observed daily oscillation in region −1185–−1224 bp in the normal group (p < 0.01) and the CO-treated group (p < 0.05) but not in the control group (Figure 6B). In the third examined region, −2000–−2054 bp, we observed the oscillation of the total methylation level only in the normal group (p > 0.0001). We observed an increased methylation level in the day in the control and CO-treated groups (p < 0.0001) for both groups compared to the normal group, while, at night, we observed increased levels in the control group (p < 0.05) and CO-treated groups (p < 0.001) compared to the normal group (Figure 6C). In the last examined region −2402–−2464 bp, we did not observe any differences (Figure 6D).
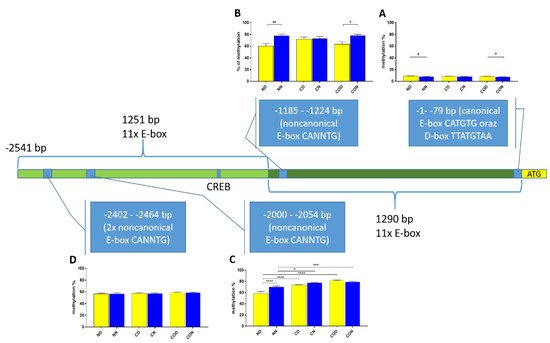
Figure 6. Scheme of promoter sequence −2500 bp downstream ATG with changes in daily total methylation level of the examined sites, −1–−79 bp (A), −1185–−1224 bp (B), −2000–−2054 bp (C), −2402–−2464 bp, (D) in the normal (N), control (C) and CO-treated groups. The differences in total methylation level were compared in each group between day (yellow bar) and night (blue bar). The bars are represented as mean ± SEM. The asterisks indicate significant differences as follows: *—p < 0.05; **—p < 0.01; ***—p < 0.001; ****—p < 0.0001.
2.3. Functional Analysis
In functional assay with the HEK 293 cell line, promoter activity was measured with relative luminescence values in 6 h intervals. Oscillation in promoter activity was observed in both the long and short promoter sequences (Figure 7). There were differences between the expression levels in the promoter sequence from DNA obtained from tissue isolated at night and day. The activity of the promoter in tissues isolated at night had lower expression level in the normal group at 8:00 PM with a long sequence (p < 0.001) and at 2:00 PM with a short sequence. Moreover, at 2:00 p.m., the short promoter showed a higher activity in samples isolated at night in the CO-treated group (p < 0.05).
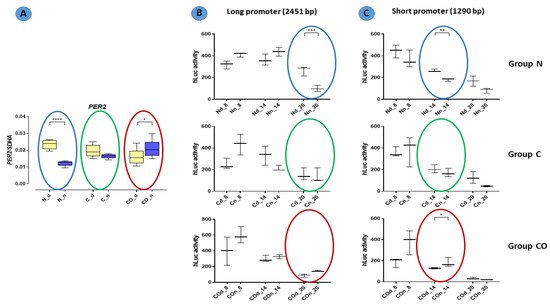
Figure 7. Promoter activity in the normal (N), control (C) and CO-treated groups. The significant differences between day (Nd, Cd, COd) and night (Nn, Cn, COn) were analyzed in each studied group. In functional assay with the HEK 293 cell line, promoter activity was measured with relative luminescence values in 6 h intervals (8 a.m., 2 p.m., 8 p.m.). Oscillation in promoter activity was observed in both the long (B) and short (C) promoter sequences. Colored circles mark the time points corresponding to the pattern expression obtained in qPCR (A) (repeated Figure 1). The asterisks indicate significant differences as follows: *—p < 0.05; **—p < 0.01; ***—p < 0.001; ****—p < 0.0001.
References
- Ripperger, J.A.; Jud, C.; Albrecht, U. The daily rhythm of mice. FEBS Lett. 2011, 585, 1384–1392.
- Welsh, D.K.; Takahashi, J.; Kay, S.A. Suprachiasmatic Nucleus: Cell Autonomy and Network Properties. Annu. Rev. Physiol. 2010, 72, 551–577.
- Takahashi, J.S. Transcriptional architecture of the mammalian circadian clock. Nat. Rev. Genet. 2017, 18, 164–179.
- Simonneaux, V.; Ribelayga, C. Generation of the Melatonin Endocrine Message in Mammals: A Review of the Complex Reg-ulation of Melatonin Synthesis by Norepinephrine, Peptides, and Other Pineal Transmitters. Pharmacol. Rev. 2003, 55, 325–395.
- Koziorowski, M.; Stefańczyk-Krzymowska, S.; Tabecka-Lonczyńska, A.; Gilun, P.; Kaminski, M. The gaseous messenger carbon monoxide is released from the eye into the ophthalmic venous blood depending on the intensity of sunlight. J. Biol. Regul. Homeost. Agents 2012, 26, 111–118.
- Oren, D.; Duda, M.; Kozioł, K.; Romerowicz-Misielak, M.; Koziorowska, A.; Sołek, P.; Nowak, S.; Kulpa, M.; Koziorowski, M. Retinal venous blood carbon monoxide response to bright light in male pigs: A preliminary study. J. Photochem. Photobiol. B 2017, 168, 12–15.
- Nowak, S.; Romerowicz-Misielak, M.; Kozioł, K.; Koziorowski, M. The seasonal changes of the heme oxygenase in the retina pig. Exp. Eye Res. 2020, 190, 107870.
- Gilun, P.; Stefanczyk-Krzymowska, S.; Romerowicz-Misielak, M.; Tabecka-Lonczynska, A.; Przekop, F.; Koziorowski, M. Carbon monoxide-mediated humoral pathway for the transmission of light signal to the hypothalamus. J. Physiol. Pharmacol. 2013, 64, 761–772.
- Sujino, M.; Nagano, M.; Fujioka, A.; Shigeyoshi, Y.; Inouye, S.-I.T. Temporal profile of circadian clock gene expression in a transplanted suprachiasmatic nucleus and peripheral tissues. Eur. J. Neurosci. 2007, 26, 2731–2738.
- Ripperger, J.A.; Albrecht, U. The circadian clock component PERIOD2: From molecular to cerebral functions. Prog. Brain Res. 2012, 199, 233–245.
- Kim, M.; De La Peña, J.B.; Cheong, J.H.; Kim, H.J. Neurobiological Functions of the Period Circadian Clock 2 Gene, Per2. Biomol. Ther. 2018, 26, 358–367.
- Bose, S.; Boockfor, F.R. Episodes of Prolactin Gene Expression in GH3 Cells Are Dependent on Selective Promoter Binding of Multiple Circadian Elements. Endocrinology 2010, 151, 2287–2296.
- Blackwell, T.K.; Weintraub, H. Differences and similarities in DNA-binding preferences of MyoD and E2A protein com-plexes revealed by binding site selection. Science 1990, 250, 1104–1110.
- Jolley, C.C.; Ukai-Tadenuma, M.; Perrin, D.; Ueda, H.R. A Mammalian Circadian Clock Model Incorporating Daytime Expression Elements. Biophys. J. 2014, 107, 1462–1473.
- Koyanagi, S.; Hamdan, A.M.; Horiguchi, M.; Kusunose, N.; Okamoto, A.; Matsunaga, N.; Ohdo, S. cAMP-response element (CRE)-mediated transcription by activating transcription Factor-4 (ATF-4) is essential for circadian expression of the Period2 Gene. J. Biol. Chem. 2011, 286, 32416–32423.
- Dioum, E.R.; Rutter, J.; Tuckerman, J.R.; Gonzales, G.; Gilles-Gonzales, M.A.; Knight, S.L. NPAS2: A gas-responsive transcription factor. Science 2002, 298, 2385–2387.
- Gilles-Gonzalez, M.A.; Gonzalez, G. Signal transduction by heme-containing PAS-domain proteins. J. Appl. Physiol. 2004, 96, 774–783.
- Lukat-Rodgers, G.S.; Correia, C.; Botuyan, M.V.; Mer, G.; Rodgers, K.R. Heme-Based Sensing by the Mammalian Circadian Protein CLOCK. Inorg. Chem. 2010, 49, 6349–6365.
More
Information
Subjects:
Biochemistry & Molecular Biology
Contributor
MDPI registered users' name will be linked to their SciProfiles pages. To register with us, please refer to https://encyclopedia.pub/register
:
View Times:
618
Revisions:
2 times
(View History)
Update Date:
05 Aug 2021
Table of Contents



Notice
You are not a member of the advisory board for this topic. If you want to update advisory board member profile, please contact office@encyclopedia.pub.
OK
Confirm
Only members of the Encyclopedia advisory board for this topic are allowed to note entries. Would you like to become an advisory board member of the Encyclopedia?
Yes
No
${ textCharacter }/${ maxCharacter }
Submit
Cancel
Back
Comments
${ item }
|
${ item.createdUser.fullName }
${ item.createdAt }
${ item.vote }
${ item.reply }
Delete
${ reply.createdUser.fullName }
${ reply.createdAt }
${ reply.vote }
Delete
There is no reply to this comment~
${ item.replyTextCharacter }/${ item.replyMaxCharacter }
Submit
Cancel
More
No more~
There is no comment~
${ textCharacter }/${ maxCharacter }
Submit
Cancel
${ selectedItem.replyTextCharacter }/${ selectedItem.replyMaxCharacter }
Submit
Cancel
Confirm
Are you sure to Delete?
Yes
No