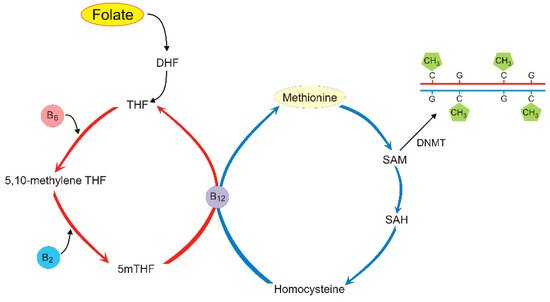
Folate (folacin, vitamin B9) is one of the B vitamins and an essential micronutrient that plays a critical role in one-carbon cellular metabolism
[46][47]. Humans, as mammals, cannot synthesize folate and must intake it with food either as a component of a natural diet, or as a fortified food or diet supplement. Folate supplementation, recommended in many countries, can come in the form of folic acid, folinic acid, or 5-methyltetrahydrofolate (5-MTHF). 5-MTHF occurs naturally and has some advantage over synthetic forms of folate including its higher bioavailability
[48]. Folate is essential for many cellular effects such as nucleoside synthesis and the methylation of biomolecules including DNA (
Figure 3).
Dietary folate is metabolized to 5-methyltetrahydrofolate (5mTHF, monoglutamyl form) by methylenetetrahydrofolate reductase (MTHFR). This reaction is important for the remethylation of homocysteine to methionine, which is a substrate for SAM, providing methyl groups for DNA methyltransferases to methylate DNA
[49]. Several other dietary nutrients are required to maintain the one-carbon flux needed for DNA methylation including vitamins B2, B6, and B12, riboflavin, and choline (
Figure 3)
[50].
Low folate status is associated with an increased risk of several disorders including cardiovascular diseases (CVD) and cancer, but the mechanisms underlying these associations are not exactly known, and several pathways may be involved
[51][52]. However, the results of some folate intervention trials suggest that excessively high folate supplementation may be detrimental for a person with an elevated risk of cancer and CVD (reviewed in
[53]). That review summarized studies with the supplementation of both folate and folic acid, which were not adequate due to inter-individual variability in the activity of the 5,10-methylene THF reductase. Therefore, the dose-effect relationship for folate in CVD may be nonlinear.
Folate deficiency could be also involved in disorders of the nervous system
[54][55]. Folate is an important factor in the functioning of the blood–brain barrier and brain development
[56]. Variability of the
MTHFR gene could result in phenotypic differences: the T allele of the 677C > T polymorphism of this gene is associated with elevated levels of plasma homocysteine
[57]. An excess of homocysteine can be detrimental for vessels and result in endothelial cell injury and changes in blood properties that can be important in CVD and migraine pathogenesis
[58][59].
Novel epigenomic loci associated with dietary folate and vitamin B12 intake were identified in a large-scale epigenome-wide association study on 5841 individuals
[60]. These studies identified significant differentially methylated positions (DMPs) and regions (DMRs) in the genome, and a pathway analysis was performed on DMR annotated genes. Vitamin B12 intake was associated with 29 DMRs annotated with 48 genes. Folate intake was negatively associated with six DMPs annotated with five genes involved in cellular processes including centrosome localization, cell proliferation, and tumorigenesis. In these studies, vitamin intake was assessed on the basis of a questionnaire.
That work can be considered in the context of the study by Illingworth et al., which assayed 1.9 million CpG islands in each of the 43 brain samples and showed over 16,000 DMRs
[61]. These authors concluded that except for the cerebellum, patterns of DNA methylation in different brain regions were more similar than the patterns for those regions in different individuals. Therefore, human brain methylome is primarily determined by DNA sequence and not developmental status. Although it is assumed and supported by many studies that the DNA methylation pattern is stable and retains in isolated genomic DNA, it is not completely known as to which changes in the epigenome are associated with death.
The 677C > T polymorphism of the
MTHFR gene is likely the most frequently addressed genetic aspect of migraine pathophysiology, but the results obtained so far are not conclusive
[62]. This polymorphism is claimed to be both an independent and combined marker for migraines, especially MA. Several meta-analyses addressing this polymorphism in migraines have been performed. Liu et al. concluded that the 677T allele was associated with an increased risk of total migraine and MA in Asians
[63]. Similar results were obtained in other analyses with the general conclusions supporting the use of folate in migraine patients, especially those with auras, but further replication studies are needed, particularly large randomized clinical trials
[62].
Menon et al. observed an inverted relationship between folic acid consumption and migraine frequency in 141 females
[64]. This relationship was modulated by the 677C > T polymorphism of the
MTHFR gene. Similar effects were noted in children with migraines and hyperhomocysteinemia
[65]. Vitamin supplementation including 2 mg/day of folic acid reduced the prevalence of MA disability from 60% to 30% after six months
[66]. A randomized, double-bind, placebo-controlled study (
n = 95) showed that folic acid at 5 mg and vitamin B6 at 80 mg decreased headache frequency and headache severity
[67].
A case-control study performed on 124 migraine patients and 130 non-migraine subjects revealed a lower level of dietary folate intake in migraineurs
[68].
No association was found between the 134R > K and 653R > Q polymorphisms of the
MTHFD1 (methylenetetrahydrofolate dehydrogenase, cyclohydrolase, and formyltetrahydrofolate synthetase 1) gene, whose product is important in folate metabolism and migraine occurrence in 162 MO and 358 MA Australian patients
[69]. Moreover, these two polymorphisms did not change the increased migraine risk associated with the 677T allele of the
MTHFR gene.
Although the exact mechanism connecting the 677C > T polymorphism of the
MTHFR gene with migraine pathophysiology is not completely known, some pathways can be considered. The C→T transition at 677 leads to the substitution of alanine to valine, thereby resulting in reduced activity of the MTHFR enzyme compared to its wild-type counterpart
[70]. Consequently, individuals homozygous for the T variant have higher homocysteine levels than the C homozygotes
[71]. As stated previously, an excess of homocysteine can be destructive for vessels and play a role in migraine pathogenesis, especially in migraine with an aura
[72]. However, the direct link between homocysteine level and migraines is still a matter of debate, especially since only one study so far has evaluated the level of homocysteine in the cerebral fluid of migraineurs
[73]. Nevertheless, elevated homocysteine may cause injury to endothelial cells, reduced flexibility of the vessels, and changes in hemostasis, which may contribute to headaches and the many associated effects and even vascular comorbidity of migraines, especially MA
[74]. Homocysteine and its related compounds may act as excitatory agonists of the NMDA (
N-methyl-
d-aspartate) subtype of glutamate receptors, which are important for CSD
[75][76]. Other potential aspects of the significance of the 677T > C polymorphism in the
MTHFR gene such as its association with calcitonin gene related peptide or migraine triggers have not been investigated so far and require further research. Several cross-sectional, prospective, or interventional studies suggest that elevated plasma levels of homocysteine are associated with an increased risk of migraines (reviewed in
[77]). The production of homocysteine requires folate and vitamins B6 and B12 whose deficiency results in DNA hypomethylation, which was hypothesized to trigger migraine resulting from an interplay with MTHFR and variants of estrogen receptor 1
[78].
In summary, folate is essential for DNA methylation and its presence in the diet was reported to exert a beneficial effect on migraines. However, these profitable effects of dietary folate have not been attributed to changes in DNA methylation or other alterations in the epigenetic profile.
 +1 point
+1 point