 +1 credit
+1 credit
| Version | Summary | Created by | Modification | Content Size | Created at | Operation |
|---|---|---|---|---|---|---|
| 1 | Guaraci Cordeiro | + 2352 word(s) | 2352 | 2021-05-31 05:09:48 | | | |
| 2 | Peter Tang | Meta information modification | 2352 | 2021-07-29 03:46:41 | | |
Video Upload Options
Bees are typically diurnal but around 1% of described species have nocturnal activity. Nocturnal bees are still poorly studied due to bias towards studying diurnal insects. However, knowledge concerning their biology and role as crop pollinators has increased. We review the literature on nocturnal bees’ traits and their host plants, and assess the crop pollination effectiveness of this neglected group. Nocturnal bees have visual adaptations to cope with low light intensities, and floral scents are a key sensory cue used to find their host flowers. Nocturnal bees generally show high flower constancy, the ability to vibrate flowers, and high transfer rates of pollen grains to stigmas. The flowers visited by nocturnal bees range from small radial and zygomorphic flowers to large brush blossoms; moreover, they visit plants with different flowering strategies. Nocturnal bees are effective pollinators of regional fruit crops in Brazil, such as cambuci (Campomanesia phaea), guaraná (Paullinia cupana), cajá (Spondias mombin), and in North America of cultivated pumpkins (Cucurbita species). However, they most likely are pollinators of several other crops. Strategies to host high numbers of nocturnal bees around cropping areas should be taken, such as preserving adjacent native forests, restricting soil management, providing food resources beyond crop flowers, and avoiding light pollution.
1. Introduction

2. How Do Nocturnal Bees Find Flowers in the Darkness?
3. Traits of Nocturnal Bees and Their Host Plants
4. Host Plants of Nocturnal Bees
5. Nocturnal Bees as Crop Pollinators
|
Plant Family |
Plant Species (Popular Name) |
Nocturnal Bee Genera |
Role as Pollinators |
Occurrence |
Ref. |
|---|---|---|---|---|---|
|
Anacardiaceae |
Spondias mombin (cajá, yellow mombin) |
Megalopta, Ptiloglossa |
E |
Caatinga- Brazil |
[35] |
|
Spondias pinnata (wild mango) |
Xylocopa |
P |
Tropical Forest-Southeast Asia |
[78] |
|
|
Cucurbitaceae |
Cucurbita foetidissima (pumpkin) |
Peponapis, Xenoglossa |
E |
Dry environments-Mexico/USA |
[79] |
|
Cucurbita maxima (pumpkin) |
Peponapis, Xenoglossa |
E |
Dry environments-Mexico/USA |
[80] |
|
|
Cucurbita pepo (pumpkin) |
Peponapis, Xenoglossa |
E |
Dry environments-Mexico/USA |
||
|
Myrtaceae |
Campomanesia phaea (cambuci) |
Megalopta, Ptiloglossa, Megommation, Zikanapis |
E |
Atlantic Forest- Brazil |
[18] |
|
Campomanesia pubescens (gabiroba) |
Megalopta, Ptiloglossa |
E |
Atlantic Forest, Cerrado-Brazil |
[81] |
|
|
Eugenia brasiliensis (grumixama) |
Megommation |
P |
Atlantic Forest- Brazil |
[39] |
|
|
Eugenia dysenterica (cagaita) |
Ptiloglossa |
P |
Cerrado-Brazil |
[39] |
|
|
Eugenia florida (guamirim) |
Megalopta |
E |
Amazon, Atlantic Forest, Caatinga, Cerrado-Brazil |
[59] |
|
|
Eugenia involucrata (cereja-do-Rio-Grande) |
Megalopta, Megommation |
P |
Atlantic Forest- Brazil |
[39] |
|
|
Eugenia neonitida (pitangatuba) |
Ptiloglossa |
P |
Atlantic Forest- Brazil |
[38] |
|
|
Eugenia punicifolia (cereja-do-cerrado) |
Ptiloglossa |
P |
Amazon, Atlantic Forest, Caatinga, Cerrado-Brazil |
[38] |
|
|
Eugenia pyriformis (uvaia) |
Ptiloglossa |
P |
Atlantic Forest- Brazil |
[39] |
|
|
Eugenia rotundifolia (abajurú) |
Ptiloglossa |
P |
Atlantic Forest- Brazil |
[38] |
|
|
Eugenia stipitata (araçá-boi) |
Megalopta, Megommation |
P |
Amazon, Atlantic Forest-Brazil |
||
|
Eugenia uniflora (pitanga) |
Ptiloglossa |
P |
Atlantic Forest- Brazil |
||
|
Myrciaria floribunda (cambuí) |
Megalopta |
E |
Amazon, Atlantic Forest, Caatinga, Cerrado-Brazil |
[59] |
|
|
Myrciaria dubia (camu-camu) |
Megalopta |
P |
Amazon-Brazil |
[83] |
|
|
Plinia cauliflora (jabuticaba) |
Ptiloglossa |
P |
Atlantic Forest, Cerrado-Brazil |
[39] |
|
|
Psidium acutangulum (araçá-pera) |
Megalopta |
P |
Amazon-Brazil |
[84] |
|
|
Syzygium malaccense (jambo-rosa) |
Ptiloglossa |
P |
Atlantic Forest- Brazil |
[39] |
|
|
Sapindaceae |
Paullinia cupana (guaraná) |
Megalopta, Ptiloglossa |
E |
Amazon-Brazil |
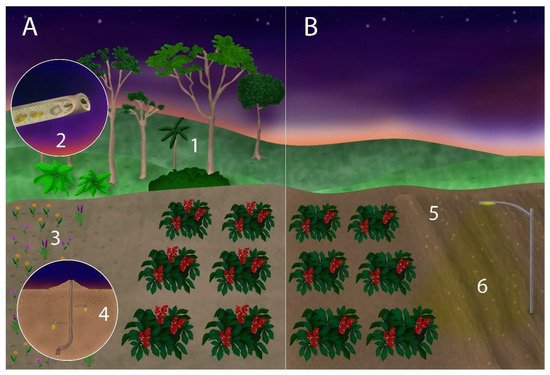
6. Requirements on Crop Areas to Host High Numbers of Nocturnal Bees
References
- United Nations. Open Working Group Proposal for Sustainable Development Goals; Full Report A/68/970; Open Working Group of the General Assembly on Sustainable Development Goals, United Nations: Geneva, Switzerland, 2014.
- Potts, S.G.; Imperatriz-Fonseca, V.L.; Ngo, H.T.; Aizen, M.A.; Biesmeijer, J.C.; Breeze, T.D.; Dicks, L.V.; Garibaldi, L.A.; Hill, R.; Settele, J.; et al. Safeguarding pollinators and their values to human well-being. Nature 2016, 540, 220–229.
- Klein, A.; Vaissiere, B.E.; Cane, J.H.; Steffan-Dewenter, I.; Cunningham, S.A.; Kremen, C.; Tscharntke, T. Importance of pollinators in changing landscapes for world crops. Proc. R. Soc. Lond. 2007, 274, 303–313.
- Aizen, M.A.; Harder, L.D. The global stock of domesticated honeybees is growing slower than agricultural demand for pollination. Curr. Biol. 2009, 19, 915–918.
- Freitas, B.; Paxton, R.A. Comparison of two pollinators: The introduced honey bee Apis mellifera and an indigenous bee Centris tarsata on Cashew Anacardium occidentale in its native range of NE Brazil. J. Appl. Ecol. 1998, 35, 109–121.
- Oliveira, R.; Schlindwein, C. Searching for a manageable pollinator for acerola orchards: The solitary oil-collecting bee Centris analis (Hymenoptera: Apidae: Centridini). J. Econ. Entomol. 2009, 102, 265–273.
- Giannini, T.C.; Alves, D.A.; Alves, R.; Cordeiro, G.D.; Campbell, A.J.; Awade, M.; Bento, J.M.S.; Saraiva, A.M.; Imperatriz-Fonseca, V.L. Unveiling the contribution of bee pollinators to Brazilian crops with implications for bee management. Apidologie 2020, 51, 406–421.
- Garibaldi, L.A.; Steffan-Dewenter, I.; Winfree, R.; Aizen, M.A.; Bommarco, R.; Cunningham, S.A.; Kremen, C.; Carvalheiro, L.G.; Harder, L.D.; Afik, O.; et al. Wild pollinators enhance fruit set of crops regardless of honeybee abundance. Science 2013, 339, 1608–1611.
- Woodcock, B.A.; Garratt, M.P.D.; Powney, G.D.; Shaw, R.F.; Osborne, J.L.; Soroka, J.; Lindström, S.A.M.; Stanley, D.; Ouvrard, P.; Edwards, M.E.; et al. Meta-analysis reveals that pollinator functional diversity and abundance enhance crop pollination and yield. Nat. Commun. 2019, 10, 1481.
- Warrant, E.J.; Kelber, A.; Gislén, A.; Greiner, B.; Ribi, W.; Wcislo, W.T. Nocturnal vision and landmark orientation in a tropical halictid bee. Curr. Biol. 2004, 14, 1309–1318.
- Kelber, A.; Warrant, E.J.; Pfaff, M.; Wallén, R.; Theobald, J.C.; Wcislo, W.T.; Raguso, R.A. Light intensity limits foraging activity in nocturnal and crepuscular bees. Behav. Ecol. 2006, 17, 63–72.
- Warrant, E.J. Nocturnal bees. Curr. Biol. 2007, 17, 991–992.
- Warrant, E.J. Seeing in the dark: Vision and visual behaviour in nocturnal bees and wasps. J. Exp. Biol. 2008, 211, 1737–1746.
- Somanathan, H.; Borges, R.M.; Warrant, E.J.; Kelber, A. Visual ecology of Indian carpenter bees I: Light intensities and flight activity. J. Comp. Physiol. A 2008, 194, 97–107.
- Wcislo, W.T.; Tierney, S.M. Behavioural environments and niche construction: The evolution of dim-light foraging in bees. Biol. Rev. 2009, 84, 19–37.
- Wcislo, W.T.; Arneson, L.; Roesch, K.; Gonzalez, V.; Smith, A.; Fernandez, H. The evolution of nocturnal behavior in sweat bees, Megalopta genalis and M. ecuadoria (Hymenoptera: Halictidae): An escape from competitors and enemies? Biol. J. Linn. Soc. 2004, 83, 377–387.
- Smith, A.R.; Kitchen, S.M.; Toney, R.M.; Ziegler, C. Is nocturnal foraging in a tropical bee an escape from interference competition? J. Insect Sci. 2017, 17, 62.
- Cordeiro, G.D.; Pinheiro, M.; Dötterl, S.; Alves-dos-Santos, I. Pollination of Campomanesia phaea (Myrtaceae) by night-active bees: A new nocturnal pollination system mediated by floral scent. Plant Biol. 2017, 19, 132–139.
- Roulston, T.H. Hourly capture rates of two species of Megalopta (Hymenoptera: Apoidea; Halictidae) at black lights in Panama with notes on nocturnal foraging by bees. J. Kans. Entomol. Soc. 1997, 70, 189–196.
- Araujo, F.F.; Araújo, P.C.S.; Siqueira, E.; Alves-dos-Santos, I.; Oliveira, R.; Dötterl, S.; Schlindwein, C. Nocturnal bees exploit but do not pollinate flowers of a common bat-pollinated tree. Arthropod Plant Interact. 2020, 14, 785–797.
- Ascher, J.S.; Pickering, J. Discover Life Bee Species Guide and World Checklist (Hymenoptera: Apoidea: Anthophila). Available online: (accessed on 31 March 2021).
- Moure, J.S. Augochlorini Beebe. In Catalogue of Bees (Hymenoptera, Apoidea) in the Neotropical Region Online Version; Moure, J.S., Urban, D., Melo, G.A.R., Eds.; Sociedade Brasileira de Entomologia: Curitiba, Brazil, 2007.
- Santos, L.M.; Melo, G.A.R. Updating the taxonomy of the bee genus Megalopta (Hymenoptera: Apidae, Augochlorini) including revision of the Brazilian species. J. Nat. Hist. 2015, 49, 575–674.
- Wcislo, W.T.; Gonzalez, V.H. Social and ecological contexts of trophallaxis in facultatively social sweat bees, Megalopta genalis and M. ecuadoria (Hymenoptera, Halictidae). Insectes Soc. 2006, 53, 220–225.
- Tierney, S.M.; Gonzales-Ojeda, T.; Wcislo, W.T. Biology of a nocturnal bee, Megalopta atra (Hymenoptera: Halictidae; Augochlorini), from the Panamanian highlands. J. Nat. Hist. 2008, 42, 1841–1847.
- Santos, L.M.; Tierney, S.M.; Wcislo, W.T. Nest descriptions of Megalopta aegis (Vachal) and M. guimaraesi Santos & Silveira (Hymenoptera, Halictidae) from the Brazilian Cerrado. Rev. Bras. Entomol. 2010, 54, 332–334.
- Urban, D.; Moure, J.S.; Melo, G.A.R. Diphaglossini Vachal. In Catalogue of Bees (Hymenoptera, Apoidea) in the Neotropical Region Online Version; Moure, J.S., Urban, D., Melo, G.A.R., Eds.; Sociedade Brasileira de Entomologia: Curitiba, Brazil, 2007.
- Roberts, R.B. Biology of the crepuscular bee Ptiloglossa guinnae n. sp. with notes on associated bees, mites and yeasts. J. Kans. Entomol. Soc. 1971, 44, 283–294.
- Rozen, J.G. Nesting biology of Diphaglossine bees (Hymenoptera, Colletidae). Am. Mus. Novit. 1984, 2786, 1–33.
- Sarzetti, L.C.; Genise, F.F.; Sánchez, V.M.; Farina, J.L.; Molina, A.M. Nesting behavior and ecological preferences of five Diphaglossinae species (Hymenoptera, Apoidea, Colletidae) from Argentina and Chile. J. Hymenop. Res. 2013, 33, 63–82.
- Keasar, T. Large carpenter bees as agricultural pollinators. Psyche 2010, 927463, 1–7.
- Michener, C.D. The Bees of the World, 2nd ed.; Johns Hopkins University Press: Baltimore, MD, USA, 2007; p. 953.
- Soares, N.C.; Morellato, L.P.C. Crepuscular pollination and reproductive ecology of Trembleya laniflora (Melastomataceae), an endemic species in mountain rupestrian grasslands. Flora 2018, 238, 138–147.
- Smith, A.R.; López-Quintero, I.J.; Moreno-Patiño, J.E.; Roubik, D.W.; Wcislo, W.T. Pollen use by Megalopta sweat bees in relation to resource availability in a tropical forest. Ecol. Entomol. 2012, 37, 309–317.
- Carneiro, L.; Martins, C.F. Africanized honeybees pollinate and preempt the pollen of Spondias mombin (Anacardiaceae) flowers. Apidologie 2012, 43, 474–486.
- Krug, C.; Cordeiro, G.D.; Schäffler, I.; Silva, C.I.; Oliveira, R.; Schlindwein, C.; Dötterl, S.; Alves-dos-Santos, I. Nocturnal bee pollinators are attracted to Guarana flowers by their scents. Front. Plant Sci. 2018, 9, 1072.
- Krug, C.; Garcia, M.V.B.; Gomes, F.B. A scientific note on new insights in the pollination of guarana (Paullinia cupana var. sorbilis). Apidologie 2015, 46, 164–166.
- Silva, A.L.G.; Pinheiro, M.C.B. Biologia floral e da polinização de quatro espécies de Eugenia L. (Myrtaceae). Acta Bot. Bras. 2007, 21, 235–247.
- Cordeiro, G.D.; Santos, I.G.F.; Silva, C.I.; Schlindwein, C.; Alves-dos-Santos, I.; Dötterl, S. Nocturnal floral scent profiles of Myrtaceae fruit crops. Phytochemistry 2019, 162, 193–198.
- Dötterl, S.; Vereecken, N. The chemical ecology and evolution of bee–flower interactions: A review and perspectives. Can. J. Zool. 2010, 88, 668–697.
- Milet-Pinheiro, P.; Ayasse, M.; Schlindwein, C.; Dobson, H.E.; Dötterl, S. Host location by visual and olfactory floral cues in an oligolectic bee: Innate and learned behavior. Behav. Ecol. 2012, 23, 531–538.
- Rachersberger, M.; Cordeiro, G.D.; Schäffler, I.; Dötterl, S. Honeybee pollinators use visual and floral scent cues to find Apple (Malus domestica) flowers. J. Agric. Food Chem. 2019, 67, 13221–13227.
- Warrant, E.J.; Kelber, A.; Wallén, R.; Wcislo, W. Ocellar optics in nocturnal and diurnal bees and wasps. Arthropod Struct. Dev. 2006, 35, 293–305.
- Greiner, B.; Ribi, W.A.; Warrant, E.J. Retinal and optical adaptations for nocturnal vision in the halictid bee Megalopta genalis. Cell Tissue Res. 2004, 316, 377–390.
- Somanathan, H.; Warrant, E.J.; Borges, R.M.; Wallén, R.; Kelber, A. Resolution and sensitivity of the eyes of the Asian honeybees Apis florea, Apis cerana and Apis dorsata. J. Exp. Biol. 2009, 212, 2448–2453.
- Jander, U.; Jander, R. Allometry and resolution of bee eyes (Apoidea). Arthropod Struct. Dev. 2002, 30, 179–193.
- Greiner, B.; Ribi, W.A.; Wcislo, W.T.; Warrant, E.J. Neural organization in the first optic ganglion of the nocturnal bee Megalopta genalis. Cell Tissue Res. 2004, 318, 429–437.
- Somanathan, H.; Kelber, A.; Borges, R.M.; Wallén, R.; Warrant, E.J. Visual ecology of Indian carpenter bees II: Adaptations of eyes and ocelli to nocturnal and diurnal lifestyles. J. Comp. Physiol. A 2009, 195, 571–583.
- Berry, R.P.; Wcislo, W.T.; Warrant, E.J. Ocellar adaptations for dim light vision in a nocturnal bee. J. Exp. Biol. 2011, 214, 1283–1293.
- Frederiksen, R.; Wcislo, W.T.; Warrant, E.J. Visual reliability and information rate in the retina of a nocturnal bee. Curr. Biol. 2008, 18, 349–353.
- Warrant, E.J. Seeing better at night: Life style, eye design and the optimum strategy of spatial and temporal summation. Vis. Res. 1999, 39, 1611–1630.
- Greiner, B.; Ribi, W.A.; Warrant, E.J. A neural network to improve dim-light vision? Dendritic fields of first-order interneurons in the nocturnal bee Megalopta genalis. Cell Tissue Res. 2005, 323, 313–320.
- Liporoni, R.; Cordeiro, G.D.; Prado, P.I.; Schlindwein, C.; Warrant, E.J.; Alves-dos-Santos, I. Light intensity regulates flower visitation in Neotropical nocturnal bees. Sci. Rep. 2020, 10, 15333.
- Kelber, A.; Roth, L.S.V. Nocturnal colour vision—Not as rare as we might think. J. Exp. Biol. 2006, 209, 781–788.
- Borges, R.M.; Somanathan, H.; Kelber, A. Patterns and processes in nocturnal and crepuscular pollination services. Q. Rev. Biol. 2016, 91, 389–418.
- Siqueira, E.; Oliveira, R.; Dötterl, S.; Cordeiro, G.D.; Alves-dos-Santos, I.; Mota, T.; Schlindwein, C. Pollination of Machaerium opacum (Fabaceae) by nocturnal and diurnal bees. Arthropod Plant Interact. 2018, 12, 633–645.
- Knudsen, J.T.; Eriksson, R.; Gershenzon, J.; Ståhl, B. Diversity and distribution of floral scent. Bot. Rev. 2006, 72, 1–120.
- Alves-dos-Santos, I.; Silva, C.I.; Pinheiro, M.; Kleinert, A.M.P. Quando um visitante floral é um polinizador? Rodriguésia 2016, 67, 295–307.
- Souza, M.A.D. Biologia Reprodutiva de Onze Espécies de Myrtaceae em Floresta de Terra Firme na Amazônia Central. Master’s Thesis, Instituto Nacional de Pesquisas da Amazônia/Universidade Federal do Amazonas, Manaus, Brazil, 1996.
- Buchmann, S.L. Buzz Pollination in Angiosperms. In Handbook of Experimental Pollination Biology; Jones, C.E., Little, R.J., Eds.; Van Nostrand & Reinhold: New York, NY, USA, 1983; pp. 73–113.
- Oliveira, F.S.; Ribeiro, M.H.M.; Nunez, C.V.; Albuquerque, P.M.C. Flowering phenology of Mouriri guianensis (Melastomataceae) and its interaction with the crepuscular bee Megalopta amoena (Halictidae) in the restinga of Lençóis Maranhenses National Park, Brazil. Acta Amaz. 2016, 46, 281–290.
- Franco, E.L.; Gimenes, M. Pollination of Cambessedesia wurdackii in Brazilian campo rupestre vegetation, with special reference to crepuscular bees. J. Insect Sci. 2011, 11, 1–13.
- Proença, C.E.; Gibbs, P.E. Reproductive biology of eight sympatric Myrtaceae from Central Brazil. New Phyt. 1994, 126, 343–354.
- Faria, F.S.; Stehmann, J.R. Biologia reprodutiva de Passiflora capsularis L. e P. pohlii Mast. (Decaloba, Passifloraceae). Acta Bot. Bras. 2010, 24, 262–269.
- Cane, J.H.; Sampson, B.J.; Miller, S.A. Pollination value of male bees: The specialist bee Peponapis pruinosa (Apidae) at summer squash (Cucurbita pepo). Environ. Entomol. 2011, 40, 614–620.
- Minter, L.M.; Bessin, R.T. Evaluation of native bees as pollinators of cucurbit crops under floating row covers. Environ. Entomol. 2014, 43, 1354–1363.
- Janzen, D. Notes on nesting and foraging behavior of Megalopta (Hymenoptera: Halictidae) in Costa Rica. J. Kans. Entomol. Soc. 1968, 41, 342–350.
- Mori, S.A.; Boeke, J.D. Pollination. In The Lecythidaceae of a Lowland Neotropical Forest: La Fumée Mountain, French Guiana; Mori, S.A., Ed.; Memoirs of the New York Botanical Garden: New York, NY, USA, 1987; pp. 137–155.
- Hopkins, M.J.G.; Hopkins, H.C.F.; Sothers, C. A Nocturnal pollination of Parkia velutina by Megalopta bees in Amazonia and its possible significance in the evolution of chiropterophily. J. Trop. Ecol. 2000, 16, 733–746.
- Schlising, R.A. Sequence and timing of bee foraging in flowers of Ipomoea and Aniseia (Convolvulaceae). Ecology 1970, 51, 1961–1967.
- Bullock, S.H.; Ayala, R.; Baker, I.; Baker, H.G. Reproductive biology of the tree Ipomoea wolcottiana (Convolvulaceae). Madroño 1987, 34, 304–314.
- Linsley, E.G.; Cazier, M.A. Some competitive relationships among matinal and late afternoon foraging activities of caupolicanine bees in southeastern Arizona. J. Kans. Entomol. Soc. 1970, 43, 251–261.
- Shelly, R.T.E.; Villalobos, E.M.; Buchmann, S.L.; Cane, J.H. Temporal patterns of floral visitation for two bee species foraging on Solanum. J. Kans. Entomol. Soc. 1993, 66, 319–327.
- Piechowski, D.; Dötterl, S.; Gottsberger, G. Pollination biology and floral scent chemistry of the Neotropical chiropterophilous Parkia pendula. Plant Biol. 2010, 12, 172–182.
- Sazima, M.; Buzato, S.; Sazima, I. Cayaponia Cabocla (Cucurbitaceae) Parece Uma Espécie Quiropterófila. In Resumos do XLVII Congresso Nacional de Botânica; Nova Friburgo: Rio de Janeiro, Brasil, 1996; pp. 407–408.
- Somanathan, H.; Krishna, S.; Jos, E.M.; Gowda, V.; Kelber, A.; Borges, R.M. Nocturnal bees feed on diurnal leftovers and pay the price of day-night lifestyle transition. Front. Ecol. Evol. 2020, 8, 566964.
- Lorenzi, H. Brazilian Trees: A Guide to the Identification and Cultivation of Brazilian Native Trees; Instituto Plantarum de Estudos da Flora: Nova Odessa, Brazil, 2002; p. 384.
- Burgett, M.; Sukumalanand, P.; Vorwohl, G. Pollen species resources for Xylocopa (Nyctomelitta) tranquebarica (F.), a night-flying carpenter bee (Hymenoptera: Apidae) of Southeast Asia. Sci. Asia 2005, 31, 65–68.
- Ordway, E.; Buchmann, S.L.; Kuehl, R.O.; Shipman, C.W. Pollen dispersal in Cucurbita foetidissima (Cucurbitaceae) by bees of the genera Apis, Peponapis and Xenoglossa (Hymenoptera: Apidae, Anthophoridae). J. Kans. Entomol. Soc. 1987, 60, 489–503.
- Fronk, W.D.; Slater, J.A. Insect fauna of cucurbit flowers. J. Kans. Entomol. Soc. 1956, 29, 141–145.
- Araujo, F.F.; Araújo, P.C.S.; Schlindwein, C. Competition between native bees and introduced honey bees on flowers of Campomanesia pubescens. 2021; manuscript in preparation.
- Falcão, M.A.; Chavéz Flores, W.B.; Ferreira, S.A.N.; Clement, C.R.; Barros, M.J.B.; Brito, J.M.C.; Santos, T.C.T. Aspectos fenológicos e ecológicos do araçá-boi (Eugenia stipitata McVaugh) na Amazônia Central. I. Plantas juvenís. Acta Amaz. 1988, 18, 27–38.
- Ferreira, G.A.C. Camu-Camu (Myrciaria dubia (Kunth) McVaugh) e Seus Polinizadores: Produtividade, Diversidade e Interações na Amazônia Central, Brasil. Master’s Thesis, Instituto Nacional de Pesquisas da Amazônia/Pós-Graduação em Botânica, Manaus, Brazil, 2020.
- Falcão, M.A.; Fereira, S.A.N.; Clement, C.R.; Santos, T.C.; Souza, R.M. Crescimento e fenologia de Araçá-Pera (Psidium acutangulum DC). Acta Amaz. 1992, 22, 285–293.
- Nadia, T.L.; Machado, I.C.; Lopes, A.V. Polinização de Spondias tuberosa Arruda (Anacardiaceae) e análise da partilha de polinizadores com Ziziphus joazeiro Mart. (Rhamnaceae), espécies frutíferas e endêmicas da caatinga. Rev. Bras. Bot. 2007, 30, 89–100.
- Mitchell, J.D.; Daly, D.C. A revision of Spondias L. (Anacardiaceae) in the Neotropics. PhytoKeys 2015, 55, 1–92.
- Caetano, C.A.; Cordeiro, G.D.; Alves-dos-Santos, I. Plants visited by crepuscular-nocturnal bees. 2021. manuscript in preparation.


