The inclusion of microalgae on ruminants' diets appears to be a promising feeding strategy to enrich milk with beneficial biomolecules such as ω3 fatty acids. However, radical alterations could occur in both structure and biochemistry potential of the rumen microbiome.
1. Introduction
The livestock sector aiming to follow the global markets and consumer trends regarding functional foods has focused on several bioactive compounds
[1]. Amongst such bioactive compounds, microalgae are considered to be a novel and sustainable nutritional alternative capable of enriching ruminants’ milk with polyunsaturated fatty acids (PUFA) that are associated with consumers’ health benefits
[2][3][4]. Notably, the dietary supplementation with
Schizochytrium spp. enriched cow
[5], sheep
[6], and goat milk
[7] with docosapentaenoic (DPA) and docosahexaenoic (DHA) fatty acids (FA). Interestingly, high-fat microalgae such as
Schizochytrium spp. could also be used for their potential in reducing methane production in ruminants as reported in vitro by Fievez et al.
[8]. Supporting this set of evidence, previous data signified the inhibiting potential of
Schizochytrium spp. on methanogenic archaea adhered to feed particles
[9] or floated in the liquid of the goats’ rumen
[10].
Nevertheless, the reprogramming of the rumen microbiome by the inclusion of PUFA-rich feed supplements tends to holistically shift the core bacteriome, concealing significant risks towards rumen function and, consequently, animal performance. It has been observed that PUFAs of marine origin exert toxic effects on the cell membrane, particularly of Gram-positive bacteria
[11], resulting in severe suppression of cellulolytic growth
[12]. In this light,
Ruminococcus species are considered to be the most sensitive taxa as they were unable to grow in the presence of DHA in in vitro cultures
[12]. Thus, nutritional strategies for methane mitigation and biohydrogenation manipulation should also take into consideration the entire rumen bacteriome to ensure that fiber digestion is not considerably compromised. It is worth mentioning that the ruminants’ ability to degrade lignocellulose derived from fibrous feedstuffs appears to be of great importance classifying ruminants as the most sustainable livestock system considering the food–feed competition between humans and animals
[13].
From this perspective, the multidisciplinary approaches that include but are not limited to the enrichment of milk and meat with bioactive molecules beneficial for human health, improved environmental sustainability through methane mitigation, and nitrogen utilization with the simultaneous guaranteed feed efficiency and animal performance shaping future livestock. However, multitargeted strategies require holistic tools, too. Until recently, the study of the rumen microbiome was implicitly linked with culture-based techniques, while most of the rumen microbes cannot be cultivated in pure cultures
[14]. Therefore, these approaches are of little help when the goal is the elucidation of the relationships between community members
[14]. The advent of meta-omics techniques such as 16S rRNA sequencing provides a much broader genomic and functional perspective in rumen microbial ecology. Up to now, the rumen bacteriome has been explored using metagenomic approaches (a) in vitro in cattle and sheep rumens in response to pure DPA, DHA, and eicosapentaenoic acid (EPA) fatty acids
[15] and (b) in vivo in early life goats in response to dietary supplementation with microalgae
Schizochytrium spp.
[16]. However, limited information exists exploring the complicated interactions between rumen proteolytic, amylolytic, and cellulolytic bacteria.
Still, dependable discrimination of ruminal bacteria according to their degradation role appears to be unreliable in generating assumptions related to their utilization activity since the majority of them used to be involved in various procedures. For instance,
Butyrivibrio fibrisolvens traditionally considered to have proteolytic, cellulolytic, and pectinolytic properties are also found to be important for PUFA biohydrogenation
[12]. However, abundances of rumen microbes are not always correlated with their overall enzymatic potential or with their end products concentration. Specifically,
Fibrobacter succinogenes, a primally cellulose-degrading bacterium, was found in a significantly lower abundance in the rumen of cows that were fed a high-concentrate ration, while carboxymethyl cellulose was prone to increase
[17]. Furthermore, it has been observed that the composition rather than the absolute abundance of the methanogens in the rumen is firmly associated with CH
4 production
[18]. Thus, the bacteriome’s synergistic action on principal enzymatic activities related to feed fermentation should also be addressed for assessing the feasibility of such novel nutritional strategies in farm-scale conditions without compromising overall sustainability.
2. Experimental Design and Animals' Diets
This study continued the analytical approach initiated in previous research works
[7][9][10][19]. The experimental trial was conducted with respect to the guidelines of the European Union Directive on the protection of animals used for scientific purposes (EU 63/2010; Council of the European Union, 2010). Briefly, twenty-four 3–4-year-old dairy goats (local (Greek) × Alpine breeds) at 150 ± 10 days in milk (DIM) were grouped into four standardized subgroups (
n = six goats/treatment) based on their fat-corrected milk (FCM6%) yield, age, and body weight (BW; 47.6 ± 5.9 kg). Each group was allocated to one of the following four groups: the control group (CON) receiving a basal diet consisting of 1 kg alfalfa hay and 1 kg concentrate/goat daily and the ALG20, ALG40, and ALG60 groups which were fed the same basal diet supplemented with 20 g, 40 g, and 60 g
Schizochytrium spp./goat, respectively (
Table 1).
Schizochytrium spp. Is traded as a commercial product DHAgold (DSM Human Nutrition & Health, Heerlen, the Netherlands). Further information about ration design, microalgae inclusion, and chemical composition of the feeds was previously reported by Mavrommatis and Tsiplakou
[7].
3. Sequencing, Quality Filtering, and Validation
A total of 2,638,775 reads was generated from the total of 40 samples, with the mean of 65,969 reads per sample. After quality filtering, 2,161,216 (82%) high-quality sequences remained, with the mean of 54,030 reads per sample. The average length was observed at 403 nts (±2.3 SD), while the mean quality was reported at 37.4 (±0.1 SD). Rarefaction curves of the bacterial population at the genus taxonomic level show that all the samples reached the plateau phase; thus, any further increase in the number of sequences would not impact the number of genera detected.
The dominant bacteria species featuring great importance in rumen functions were amplified using a well-established qPCR platform, validating the NGS results. The data in both cases were transformed as fold change compared to their control groups. Amongst the groups, the same pattern was observed between the qPCR and NGS data. Only for Butyrivibrio fibrisolvens, a minor discrepancy was found between qPCR and NGS in the ALG20 group; however, in both analyses, the differences were not significant.
4. Diversity, Richness, and Composition of the Ruminal Bacteriome
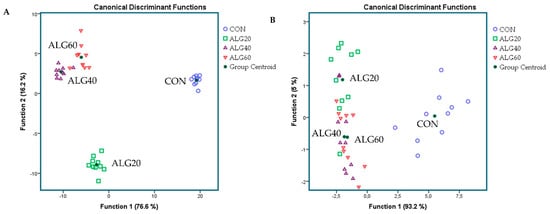
Figure 1A,B depicts the discriminant plots of the four dietary treatments (CON, blue ○; ALG20, green □; ALG40, purple △; and ALG60, red ▽) throughout the experimental period based on the forty-three most abundant genera (>0.06%) within the goats’ rumen. By inserting the variables independent-together, the proportions of the samples that were correctly classified were 100%, while Wilks’s λ was observed at < 0.001 for Function 1 (p < 0.001) and at 0.002 for Function 2 (p < 0.001) (Figure 1A). CON variables were significantly (Function 1) different from those of the microalgae-fed groups, while ALG20 was further allocated to the left bottom of the plot, indicating a dose response between 20 and 40 g of Schizochytrium spp. dietary supplementation of the rumen bacteriome (Figure 1A). Applying a stepwise method aiming to avoid any multicollinearity bias, a lower correct classification was achieved (80%). Wilks’s λ was observed at 0.042 for Function 1 (p < 0.001) and at 0.513 for Function 2 (p = 0.012), while the abundances of Intestinimonas, Flintibacter, Anaerovibrio, Butyrivibrio, Anaerocolumna, and Endomicrobium were the variables that contributed the most (Figure 1B). CON variables were significantly (Function 1) different from those of the microalgae-fed groups, while ALG20, ALG40, and ALG60 showed a minor overlap (Function 2), indicating that there was no significant dose-dependent effect considering the stepwise method (Figure 1B).
Figure 1. Discriminant plots separating (A) the four dietary treatments (CON; blue ○, ALG20; green □, ALG40; purple △, and ALG60; red ▽) according to pooled data of two sampling times (20th and 40th experimental day) that entered independents together on the abundances explained the forty-three most abundant genera in goats’ rumen (> 0.06%) and (B) discriminating the four dietary treatments (CON; blue ○, ALG20; green □, ALG40; purple △, and ALG60; red ▽) based on a step-wise method. CON = control concentrate without microalgae (Schizochytrium spp.); ALG20 = CON with 20 g/Kg Schizochytrium spp.; ALG40 = CON with 40 g/Kg Schizochytrium spp.; ALG60 = CON with 60 g/Kg Schizochytrium spp.
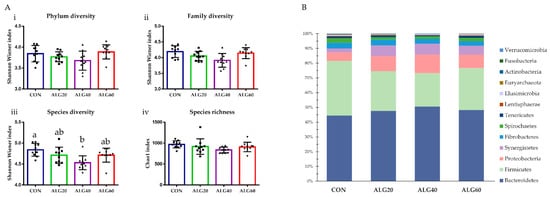
The Shannon–Wiener index did not differ between the phyla (p = 0.171), families (p = 0.055), and genera (p = 0.146), while in the ALG40 group, Shannon diversity at the species level (p = 0.033) was decreased significantly compared to the CON group (Figure 2A). Despite the significant reduction in species diversity, Chao1 richness in ALG40 was decreased only numerically (970 vs. 837; p = 0.271) (Figure 2A). The dominant phyla appeared to be Bacteroidetes (CON, 18.86%; ALG20, 21.91%; ALG40, 22.50%; ALG60, 18.92%), Firmicutes (CON, 15.66%; ALG20, 12.35%; ALG40, 10.07%; ALG60, 11.19%), and Proteobacteria, while some minor populations of Synergistetes, Fibrobacteres, Spirochaetes, Tenericutes, Lentisphaerae, Elusimicrobia, Euryarchaeota, Actinobacteria, Fusobacteria, and Verrucomicrobia were also identified (Figure 2B). Actinobacteria were significantly decreased (p = 0.021) in the ALG40 and ALG60-fed goats compared to the CON group. Bacteroidetes, the predominant rumen phylum, tended to increase (p = 0.099) by 19% in the ALG40-fed goats compared to those that consumed the control diet. Firmicutes were decreased (p = 0.012) in the microalgae-fed goats compared to the CON group. Proteobacteria were increased significantly (p = 0.019) in the ALG20- and ALG40-fed goats compared to the CON group, while the clusters unable to be annotated were significantly lower (p < 0.001) in the ALG20 and ALG40 groups compared to the CON and ALG60 ones. Synergistetes were significantly increased (p = 0.012) in the microalgae-fed goats compared to the CON group.
Figure 2. (A) Effects of supplementing with microalgae Schizochytrium spp. on (i) phylum, (ii) family, and (iii) species diversity based on the Shannon–Wiener index and (iv) Chao1 richness at the species level. Bars with different superscripts (a, b) mean dietary treatments differ significantly (p ≤ 0.05) according to the analysis of variance (ANOVA) using a general linear model for repeated measures; post hoc analysis was performed when appropriate using the Tukey’s multiple range test. (B) Relative abundances of the identified phyla (unidentified clusters and no-hits were omitted) in the four dietary treatments (CON, ALG20, ALG40, and ALG60) within the two sampling times (20th and 40th days) illustrated in the cumulative bar graph. After the exclusion of the unmapped clusters and no-hit sequences, the relative abundances of the identified phyla were transformed as the percentage of the total identified taxa. CON = control concentrate without microalgae (Schizochytrium spp.); ALG20 = CON with 20 g/Kg Schizochytrium spp.; ALG40 = CON with 40 g/Kg Schizochytrium spp.; ALG60 = CON with 60 g/Kg Schizochytrium spp.
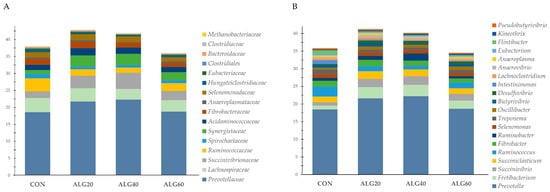
Figure 3A depicts the relative proportion of the sixteen most abundant (>0.1%) bacteria families within the goats’ rumen. Prevotellaceae, the most abundant family, tended to increase in the ALG40 group compared to the control (18.63 vs. 22.33%; p = 0.100). Succinivibrionaceae and Synergistaceae were increased significantly (p = 0.038 and p = 0.012, respectively) while Bacteroidaceae were decreased (p = 0.050) in the ALG20- and ALG40-fed goats compared to the CON. Ruminococcaceae were significantly decreased (p = 0.011) in the microalgae-fed goats compared to the normally fed ones. Eubacteriaceae, Clostridiales, Endomicrobiaceae, Erysipelotrichaceae, Atopobiaceae, and Eggerthellaceae were significantly decreased (p < 0.05) in the goats whose diets were supplemented with Schizochytrium spp., while Hungateiclostridiaceae tended to decrease (p = 0.089) in the ALG40 group compared to the CON group. The methanogenic archaea family of Methanobacteriaceae tended to decrease (p = 0.077) in the ALG40 and ALG60 goats compared to the ALG20 group.
Figure 3. Relative abundances of the most dominant (A) families (>0.1%) and (B) genera (>0.3%) in the four dietary treatments (CON, ALG20, ALG40, and ALG60) within the two sampling times (20th and 40th days) illustrated in the cumulative bar graph. CON = control concentrate without microalgae (Schizochytrium spp.); ALG20 = CON with 20 g/Kg Schizochytrium spp.; ALG40 = CON with 40 g/Kg Schizochytrium spp.; ALG60 = CON with 60 g/Kg Schizochytrium spp.
At the genus level, a total of 578 genera was annotated. Among these genera, the 47 most abundant ones were statistically assessed, and the first 20 (>0.3%) are presented as the relative proportion in Figure 3B. Following the trend of their family level, Prevotella species tended to increase in the ALG40-fed goats, while Ruminococcus were decreased significantly (p = 0.026) in the microalgae-fed goats compared to the CON goats. Fretibacterium were increased (p = 0.012), while Hungateiclostridium were decreased (p = 0.018) in the ALG20 and ALG40 rumen liquid compared to the CON. Ruminobacter species showed a significant increase (p = 0.049) in the ALG40 rumen fluid compared to the ALG60 and CON ones. Flintibacter, Intestinimonas, Eubacterium, Sporobacter, Neglecta, Blautia, Endomicrobium, and Stomatobaculum were decreased significantly (p < 0.05) in the goats whose diets were supplemented with Schizochytrium spp. compared to the normally fed goats. Anaerovibrio showed a significant increase (p = 0.049) in the ALG20 group compared to the CON and ALG60 groups. A tendency for increase (p = 0.074) was observed in Methanomicrobium in the ALG60 group compared to the other dietary treatments including the CON.
5. Relative Abundance of the Predominant Bacteria Species in the Goats’ Rumen
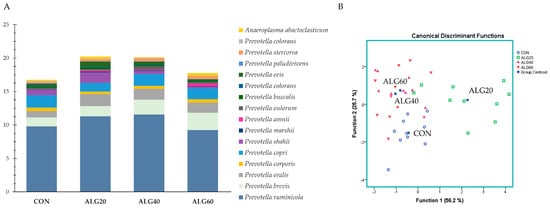
Figure 4A depicts the abundances of the goats’ rumen species featuring a pivotal role in protein, amino acid, and peptide degradation without ruling out their involvement in other functions, while Figure 4B describes those variables’ ability to correctly classify the dietary treatment. Prevotella ruminicola was found to be the dominant Bacteroidetes in the goats’ rumen. Prevotella brevis tended to increase (p = 0.097) in the ALG60 group compared to the CON. Since no considerable fluctuations were observed at the species level, only a tendency for discrimination (p = 0.098; Wilks’s λ = 0.110) was found (Figure 4B). Specifically, the ALG20 group was located in the upper right corner of the plot away from the CON group (Function 1).
Figure 4. (A) Abundances of the most representative proteolytic species in the four dietary treatments (CON, ALG20, ALG40, and ALG60) within the two sampling times (20th and 40th days) illustrated in clustered stacked columns. (B) Discriminant plots separating the four dietary treatments (CON, blue ○; ALG20, green □; ALG40, purple △; and ALG60, red ▽) according to the pooled data of the two sampling times (20th and 40th experimental days) entered together as independents on the abundances of the sixteen principal species with proteolytic activity in the goats’ rumen. CON = control concentrate without microalgae (Schizochytrium spp.); ALG20 = CON with 20 g/Kg Schizochytrium spp.; ALG40 = CON with 40 g/Kg Schizochytrium spp.; ALG60 = CON with 60 g/Kg Schizochytrium spp.
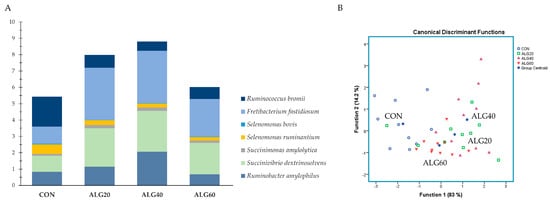
Figure 5A summarized the species with pronounced amylolytic activity. Ruminobacter amylophilus, prominent amylolytic bacteria within the rumen, were significantly increased in the ALG40-fed goats compared to the CON and ALG60 ones. Succinivibrio dextrinosolvens also showed a significant (p = 0.044) enhancement in the ALG20 and ALG40 groups, while Fretibacterium fastidiosum was increased (p = 0.012) in the microalgae-fed goats compared to the CON group. Due to the aforementioned alterations, the CON group was significantly different from the ALG20 and ALG40 groups in Function 1 (p = 0.012; Wilks’s λ = 0.327). However, a 60% correct classification was achieved.
Figure 5. (A) Abundances of the most representative amylolytic species in the four dietary treatments (CON, ALG20, ALG40, and ALG60) within the two sampling times (20th and 40th days) illustrated in clustered stacked columns. (B) Discriminant plots separating the four dietary treatments (CON, blue ○; ALG20, green □; ALG40, purple △; and ALG60, red ▽) according to the pooled data of the two sampling times (20th and 40th experimental days) entered together as independents on the abundances of the seven principal species with amylolytic activity in the goats’ rumen. CON = control concentrate without microalgae (Schizochytrium spp.); ALG20 = CON with 20 g/Kg Schizochytrium spp.; ALG40 = CON with 40 g/Kg Schizochytrium spp.; ALG60 = CON with 60 g/Kg Schizochytrium spp.
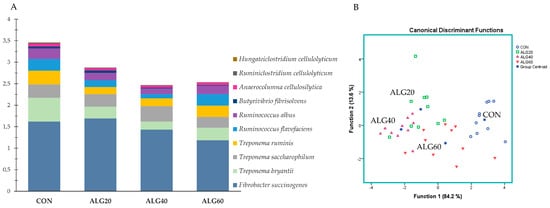
Figure 6A summarized the overall reduction of cellulolytic species in the microalgae-fed rumen fluid. The dominant cellulose degradation species, Ruminococcus flavefaciens, was decreased (p = 0.016) significantly in the ALG40 group compared to the CON and ALG60 groups. Treponema bryantii was considerably (p < 0.001) decreased in the Schizochytrium spp.-supplemented goats, while Ruminococcus gauvreauii and Ruminococcus albus tended to decrease (p = 0.070 and p = 0.099, respectively) in the ALG40 group compared to the CON. These variations resulted in significant discrimination of the dietary treatments according to the most abundant cellulolytic bacteria (Figure 6B). Specifically, the CON group was mapped on the right-hand side of the plot, clearly distanced from the ALG20 and ALG40 groups for Function 1 (p < 0.001; Wilks’s λ = 0.115). Overall, 77.4% of the variables were correctly classified, while the CON variables achieved a higher classification score (90%).
Figure 6. (A) Abundances of the most representative cellulolytic species in the four dietary treatments (CON, ALG20, ALG40, and ALG60) within the two sampling times (20th and 40th days) illustrated in clustered stacked columns. (B) Discriminant plots separating the four dietary treatments (CON, blue ○; ALG20, green □; ALG40, purple △; and ALG60, red ▽) according to the pooled data of the two sampling times (20th and 40th experimental days) entered together as independents on the abundances of the ten principal species with cellulolytic activity in the goats’ rumen. CON = control concentrate without microalgae (Schizochytrium spp.); ALG20 = CON with 20 g/Kg Schizochytrium spp.; ALG40 = CON with 40 g/Kg Schizochytrium spp.; ALG60 = CON with 60 g/Kg Schizochytrium spp.
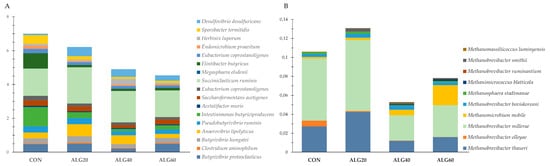
Figure 7A presents a group of rumen bacteria with a key role in nutrients degradation and utilization. Anaerovibrio lipolyticus were increased significantly (p = 0.049) in the ALG20-fed goats, while Intestinimonas butyriciproducens and Flintibacter butyricus were decreased (p < 0.001) in the microalgae-fed goats compared to the CON group (Figure 7A). Additionally, Endomicrobium proavitum were significantly increased in the rumen fluid of the goats fed with microalgae compared to the CON group (p = 0.009). Succiniclasticum ruminis and Herbinix luporum tended to increase in the ALG20 and ALG40 groups, respectively, compared to the CON group (p = 0.098; p = 0.068). Another important bacterium showing a significant reduction in the microalgae-fed goats was Sporobacter termitidis (p = 0.003). On the other hand, Desulfovibrio desulfuricans were significantly increased in the ALG20 and ALG40 groups compared to the CON group (p = 0.049). Figure 7B depicts the total abundance of Euryarchaeota species that were identified in the goats’ rumen. Methanobrevibacter olleyae were significantly decreased (p < 0.001) in the microalgae-fed goats compared to the CON group, while Methanomicrobium mobile tended to increase (p = 0.077) in the ALG60 group. Methanosphaera stadtmaniae, Methanobrevibacter millerae, and Methanobrevibacter thaueri decreased only numerically (p > 0.10) in the ALG40-fed goats compared to the CON group. Considering the sum of the Methanobrevibacter olleyae, Methanosphaera stadtmaniae, Methanobrevibacter millerae, and Methanobrevibacter thaueri abundances, a tendency for decrease (p = 0.095) was observed in the ALG40 goats.
Figure 7. Abundances of (A) influential rumen species, (B) and methanogenic archaea in the four dietary treatments (CON, ALG20, ALG40, and ALG60) within the two sampling times (20th and 40th days) illustrated in clustered stacked columns. CON = control concentrate without microalgae (Schizochytrium spp.); ALG20 = CON with 20 g/Kg Schizochytrium spp.; ALG40 = CON with 40 g/Kg Schizochytrium spp.; ALG60 = CON with 60 g/Kg Schizochytrium spp.
6. Enzyme Activities, Ammonia Concentration, and Ruminal pH
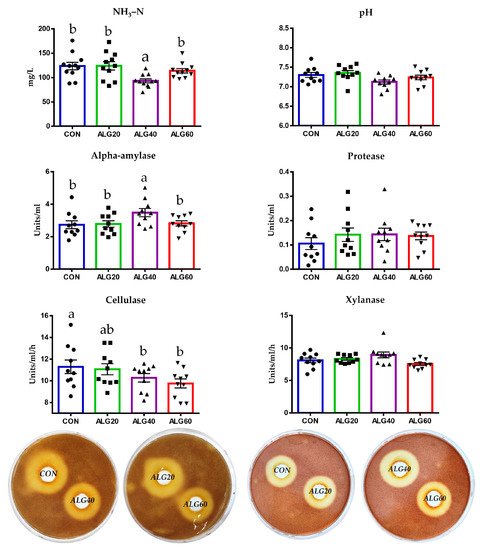
Ammonia concentration was significantly decreased in the ALG40-fed goats compared to the other groups including the CON (Figure 8).
Figure 8. Effects of supplementing with microalgae Schizochytrium spp. on pH, ammonia concentration, alpha-amylase, protease, cellulase, and xylanase activity in the rumen fluid for the four dietary treatments (CON, ALG20, ALG40, and ALG60) within the two sampling times (20th and 40th days) illustrated in bar graphs ± SEM. Alpha-amylase and protease were determined spectrophotometrically, while cellulase and xylanase activities were assayed (individually) using Petri dishes. Petri dishes depict the area of the zone of clearing of the pooled rumen fluid of each dietary treatment which runs as a preliminary trial prior to the individual ones. Bars with different superscripts (a, b) between dietary treatments differ significantly (p ≤ 0.05) according to the analysis of variance (ANOVA) using a general linear model for repeated measures and post hoc analysis was performed when appropriate using the Tukey’s multiple range test. CON = control concentrate without microalgae (Schizochytrium spp.); ALG20 = CON with 20 g/Kg Schizochytrium spp.; ALG40 = CON with 40 g/Kg Schizochytrium spp.; ALG60 = CON with 60 g/Kg Schizochytrium spp.
In detail, ammonia was determined to be 25% lower in the ALG40 group compared to the CON group (p = 0.008). Ruminal pH did not differ significantly (p > 0.05) between the dietary treatments. Alpha-amylase activity was increased in the ALG40 group compared to the other dietary treatments (Figure 8). More specifically, an increase of 27% was found in the ALG40 group compared to the CON group (p = 0.048). Protease activity portrayed an upward trend in the microalgae-fed groups; however, the enhancement was not significantly considerable (p = 0.456; Figure 8). Cellulase and xylanase activities were determined by measuring the zones of clearance of the rumen inoculum in the cellulose and xylan agar Petri dishes, respectively. Cellulase activity was decreased (p = 0.050) in the ALG40 and ALG60 groups compared to the CON group, while the ALG20-fed goats were not affected significantly. Xylanase activity was not considerably different (p = 0.356).
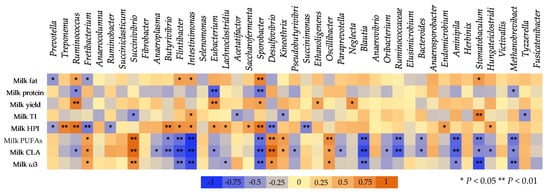
7. The Correlation between Ruminal Bacteria and Milk Composition
Figure 9 presents the Spearman’s correlation between the dominant microbial genera in the goats’ rumen with the goats’ milk performance and its pivotal constituents. Milk fat was negatively correlated (p < 0.05) with the abundance of Prevotella and Fretibacterium species, while a positive correlation was found with Ruminococcus, Flintibacter, Intestinimonas, and Sporobacter. Milk protein was negatively correlated with Ruminococcus (p < 0.05), Eubacterium (p < 0.01), Sporobacter (p < 0.01), and Methanobrevibacter (p < 0.05) abundances. Positive correlations were observed between milk yield and Ruminococcus (p < 0.01), Eubacterium (p < 0.01), Sporobacter (p < 0.05), Ethanoligenens (p < 0.05), and Neglecta (p < 0.05) abundances. The health-promoting index (HPI) which used to evaluate the potential nutritional value of milk was negatively correlated with Prevotella (p < 0.05), Fretibacterium (p < 0.01), Succinimonas (p < 0.01), and Ruminobacter species (p < 0.05), while positive correlation was observed with Treponema (p < 0.01), Ruminococcus (p < 0.01), Butyrivibrio (p < 0.01), Flintibacter (p < 0.05), Intestinimonas (p < 0.05), Eubacterium (p < 0.05), Lachnoclostridium (p < 0.05), Saccharofermenta (p < 0.05), Endomicrobium (p < 0.05), Hungateiclostridium (p < 0.05), and Sporobacter (p < 0.01).
Figure 9. Spearman’s correlation between the most abundant genera within the goats’ rumen and milk performance and its important constituents. Milk fat expressed as percentage, milk protein expressed as percentage, milk yield expressed as ml/day; milk TI (thrombogenic index) = (C14:0 + C16:0 + C18:0)/(0.5 × MUFA) + (0.5 × ω6 PUFA) + (3 × ω3 PUFA) + (ω3 PUFA/ω6 PUFA); milk HPI (health-promoting index) = (ω6 PUFA + ω3 PUFA + MUFA)/(C12:0 + 4 × C14:0 + C16:0); Milk PUFAs (polyunsaturated fatty acids) = cis-9, trans-11 C18:2 + C18:2n−6c + C18:2n−6t + C18:3n−3 + C18:3n−6 + C20:3n−3; milk CLA (cis-9, trans-11 C18:2); and milk ω3—the sum of C18:3n−3, C20:3n−3, and C22:6n−3.
Milk polyunsaturated fatty acids (PUFA) were negatively correlated with Butyrivibrio (p < 0.05), Flintibacter (p < 0.01), Intestinimonas (p < 0.01), Sporobacter (p < 0.01), Blautia (p < 0.01), Ruminococcaceae (p < 0.01), Bacteroides (p < 0.05), Aminipila (p < 0.01), Stomatobaculum (p < 0.01), and Methanobrevibacter (p < 0.01). On the contrary, positive correlation was found with Fretibacterium, Succinivibrio, Desulfovibrio, Kineothrix, and Oscillibacter. Milk conjugated linoleic acid showed negative correlation with Ruminococcus (p < 0.05), Anaeroplasma (p < 0.05), Butyrivibrio (p < 0.01), Flintibacter (p < 0.01), Intestinimonas (p < 0.01), Eubacterium (p < 0.05), Sporobacter (p < 0.01), Pseudobutyrivibrio (p < 0.05), Paraprevotella (p < 0.05), Blautia (p < 0.01), Oribacterium (p < 0.05), Ruminococcaceae (p < 0.01), Bacteroides (p < 0.05), Aminipila (p < 0.01), Hungateiclostridium (p < 0.05), and Methanobrevibacter (p < 0.05). In contrast, CLA and total milk ω3 fatty acids were positively correlated with Fretibacterium (p < 0.05), Succinivibrio (p < 0.01), Desulfovibrio (p < 0.05), and Oscillibacter (p < 0.05).
8. Conclusions
Although the cellulolytic potential was substantially suppressed, the proteolytic and amylolytic functions tended to be enhanced, indicating an adaptation mechanism in order to preserve lactation in the medium inclusion level (40 g Schizochytrium spp.). Generally, alternate beneficial modulations were observed related to functional end products and environmental impact; at the same time, warnings flags arose regarding the animal performance and balanced homeostasis. In this context, future rumen microbiome engineering should not only confirm the aforementioned observation in the metabolic spectrum (transcriptomic and proteomic approaches), but also evaluate the synergistic potential of several feed additives to counteract the arising challenges.
 +1 credit
+1 credit