Your browser does not fully support modern features. Please upgrade for a smoother experience.
Submitted Successfully!
 +1 credit
+1 credit
Thank you for your contribution! You can also upload a video entry or images related to this topic.
For video creation, please contact our Academic Video Service.
| Version | Summary | Created by | Modification | Content Size | Created at | Operation |
|---|---|---|---|---|---|---|
| 1 | Fédérico Perche | + 1990 word(s) | 1990 | 2021-06-28 08:35:20 | | | |
| 2 | Lily Guo | Meta information modification | 1990 | 2021-07-12 02:44:04 | | |
Video Upload Options
We provide professional Academic Video Service to translate complex research into visually appealing presentations. Would you like to try it?
Cite
If you have any further questions, please contact Encyclopedia Editorial Office.
Perche, F. Lipid-Based mRNA Nanoparticles. Encyclopedia. Available online: https://encyclopedia.pub/entry/11886 (accessed on 08 February 2026).
Perche F. Lipid-Based mRNA Nanoparticles. Encyclopedia. Available at: https://encyclopedia.pub/entry/11886. Accessed February 08, 2026.
Perche, Fédérico. "Lipid-Based mRNA Nanoparticles" Encyclopedia, https://encyclopedia.pub/entry/11886 (accessed February 08, 2026).
Perche, F. (2021, July 09). Lipid-Based mRNA Nanoparticles. In Encyclopedia. https://encyclopedia.pub/entry/11886
Perche, Fédérico. "Lipid-Based mRNA Nanoparticles." Encyclopedia. Web. 09 July, 2021.
Copy Citation
Messenger RNA (mRNA) is being extensively used in gene therapy and vaccination due to its safety over DNA, in the following ways: its lack of integration risk, cytoplasmic expression, and transient expression compatible with fine regulations. However, clinical applications of mRNA are limited by its fast degradation by nucleases, and the activation of detrimental immune responses. Advances in mRNA applications, with the recent approval of COVID-19 vaccines, were fueled by optimization of the mRNA sequence and the development of mRNA delivery systems. Although delivery systems and mRNA sequence optimization have been abundantly reviewed, understanding of the intracellular processing of mRNA is mandatory to improve its applications.
mRNA delivery
intracellular routing
lipid-based nanoparticles
1. Introduction
The in vivo administration of mRNA for in situ protein production was first reported in 1990, after intramuscular injection of an mRNA coding a reporter gene [1]. Since, there has been a growing interest in mRNA as a therapeutic modality, especially in the last decade [2][3]. It can be used as a platform for protein replacement therapies, genome engineering, cellular reprogramming, tolerization for allergy, and immunotherapies including vaccination [3][4]. Compared to plasmid DNA (pDNA), mRNA does not need to reach the nucleus, as it is expressed in the cytosol where the translation machinery resides. Hence, the mRNA approach is also safer, with no possibility of mutation, integration, or other undesirable genetic events [3]. In addition, it offers the possibility to transfect difficult-to-transfect differentiated cells, such as dendritic cells and neurons [5][6][7][8][9]. Moreover, mRNA is produced in vitro in a cell-free system, and methods and facilities for the large-scale production of therapeutic mRNA have been built to produce the approved SARS-CoV2 mRNA vaccines [10][11].
Since RNA molecules are unstable, prone to extracellular nucleases degradation, and are membrane-impermeable, mRNA delivery systems are required to protect them from degradation and to promote their cellular uptake into the targeted cells. Therefore, delivery systems allow them to be protected from rapid degradation in biological fluids as well as to enhance their cell uptake. In fact, it has been shown that free circulating RNA can be degraded within 15 s in the presence of blood RNAses (which are even more present in cancer patients), and mRNA has an estimated half-life of 1–2 min in human serum or bovine vitreous [12][13].
To date, different formulations have been designed to improve the stability and the intracellular delivery. Lipid-based systems represent the majority of those formulations, besides those comprising polymer, micellar or lipid–polymer hybrid [2][3][14][15][16]. In all cases, as their size ranges from a hundred nanometers to a few micrometers, these formulations are categorized as nanoparticles (NP). To deliver their cargo intracellularly, NP need to diffuse across the dense extracellular matrix and across the plasma membrane [17][18]. The main mechanism for NP to cross these barriers is endocytosis, where invaginations of the cell membrane confine NP in endosomal vesicles targeted for degradation [19]. Accordingly, NP need to incorporate features to control cell targeting, intracellular trafficking and the intracellular release of the mRNA in the cytoplasm for its translation [20]. Another important limitation to the delivery of synthetic mRNA is its recognition as foreign mRNA by intracellular sensors, such as Toll-like receptors (TLRs) 7 and 8. This recognition induces a type I interferon (INF α/β) response, which induces the suppression of mRNA translation as well as a detrimental inflammatory response [11][21]. Interestingly, in addition to mRNA, the chemical groups of the vector have been reported to be sensed by the immune system as well, as reported by Anderson’s lab [22].
The most advanced lipid-based NP for mRNA delivery are lipid nanoparticles (LNPs) (reviewed in [11][15][23]). LNPs are prepared by the microfluidic mixing of lipids in ethanol and mRNA in acidic buffer (pH ≤ 4.0). Lipids include an ionizable lipid (pKa < 7) that will be protonated at acidic pH to condense mRNA and release mRNA inside the cells, cholesterol for stabilization, and helper lipids for endosomal escape DOPE (1,2-dioleoyl-sn-glycero-3-phosphatidyl-ethanolamine) or DSPC (distearoylphosphatidylcholine) and a PEGylated (polyethylene glycol) lipid to prevent aggregation of LNPs [3]. LNPs combine the following several advantages: a high mRNA encapsulation efficiency, can be injected by various routes, preclinical and clinical proof of activity, and stability under storage (reviewed in [3][11][24]). Moreover, their production is reproducible and several techniques exist for the large-scale production of mRNA LNPs [25]. Accordingly, our review will focus on LNPs-mediated delivery of mRNA.
Despite the lack of correlation between in cellulo mRNA transfection and in vivo delivery efficacy [26][27], the evaluation of mRNA delivery systems in cell culture still represents the first step of the formulation design. Accordingly, understanding the correlation between intracellular bottlenecks and therapeutic activity is critical to further advance mRNA therapy [3][28]. There are very few studies that address the cellular uptake and subcellular distribution of exogenous mRNA [29].
The first step in mRNA delivery is to reach the targeted organ where the therapeutic mRNA should be expressed. Several units in the LNPs composition stabilize the LNPs to maximize the dose reaching the target cell. Such units should prevent sedimentation of LNPs in the storage medium, prevent aggregation in the physiological medium they are dispersed into after injection, limit drug release after interaction with the factors present in physiological fluids, and improve accumulation in the targeted organs and targeted cells. To grasp the complexity of the NP journey, we refer the reader to a review by AT Florence, describing the challenges faced by NP after administration into the body [30]. Decreasing mRNA leakage from LNPs and improving their circulation time have been the focus of extensive research (reviewed in [25]).
Indeed, after intravenous injection, most NP are captured by innate defense systems against exogenous agents [31]. Surface modification with lipids conjugated to hydrophilic PEG (polyethylene glycol) groups decreases protein adsorption onto LNPs and prevents their aggregation in the circulation [15]. As opsonization by serum-borne proteins marks NPs for elimination by cells of the mononuclear phagocyte system in the spleen and liver, PEGylation increases the circulation time of LNPs and their chance to reach the target organ [25]. The benefit of PEGylation is not limited to intravenous injection, as the decreased opsonization and neutralization of surface charge by PEG groups also favors the delivery to draining lymph nodes after intramuscular injection, which is a critical parameter for vaccination [32][33]. However, PEGylation also decreases the cellular uptake of NP and their endosomal escape ability [31]. The compromise between enough PEG-lipid to decrease non-specific interactions, yet not too much to allow endocytosis, has been referred to as the “PEG dilemma” [34]. Further, mRNA LNPs usually contain 1.5% PEGylated lipid, which is a percentage that is sufficient to avoid LNPs aggregation during storage and limit protein adsorption after infusion [11][15][23][24][35]. To resolve the PEG dilemma, ligands are also attached to a fraction of PEGylated lipids for cellular targeting. Stimuli-sensitive PEGylation, with PEGylated units activated by stimuli enriched in the target organ, are another solution to the PEG dilemma (reviewed in [36]).
The other critical parameter for the extracellular stability of LNPs is the presence of cholesterol, which is a feature already reported essential to decrease drug leakage from liposomes [37]. Cholesterol increases the viscosity of the surface and increases the encapsulation efficiency (reviewed in [25]). The presence of cholesterol impedes lipid extraction by high-density lipoproteins, which would cause LNPs breakdown and mRNA leakage. Finally, the ionizable lipid ensures the complexation of mRNA in a segregated region of LNPs, protecting it from degradation by extracellular nucleases [35][38]. In this review, we describe different approaches to study cellular uptake, the endosomal escape, and intracellular sensing of delivered mRNA (Figure 1).
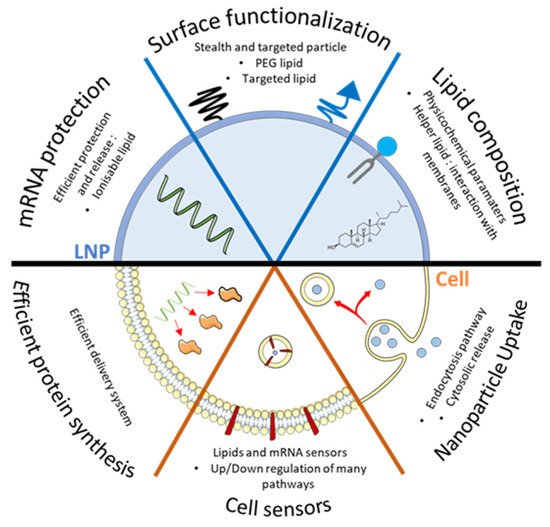
Figure 1. Critical parameter to consider regarding LNPs formulation and cellular interactions. To perform efficient transfection, LNPs should protect mRNA with ionizable lipid until its final destination and should be able to release it efficiently. Surface functionalization through incorporation of targeted lipid will help to reach specific organ/cells while PEGylated lipid will help the circulation of particles in vivo. Finally, incorporation of helper lipids will help either the formation of LNPs or its interaction with biological membrane. Altogether, those lipids need to trigger an efficient cellular uptake of the particle, which leads to the cytosol and avoids the lysosome, without triggering deleterious cell sensors, so that mRNA can lead to an efficient protein production in the cell.
2. Quick Look to Other mRNA Loaded Formulations
LNPs are part of the latest innovation in terms of a lipidic formulation to deliver mRNA, thanks to the use of microfluidic tools to produce them, but many studies involved other well-known formulations, such as liposome, micelles, or polymers. Liposomes are vesicles formed from lipids (synthetic or natural), which formed a lipid bilayer and an internal space capable of encapsulating molecules such as small drugs [39]. In 1987, Felgner and coworkers used a cationic liposome for gene therapy (with DNA) for the first time, made with DOTMA (N-[1-(2,3-dioleyloxy)propyl]-N,N,N-trimethylammonium chloride), and in 1989, the same team performed transfection with mRNA [40][41]. Complexes made of liposomes and nucleic acids are called lipoplexes; the nucleic acid is embed through the bilayer, thanks to electrostatic interactions [42], which can impact the surface aspect and stability of the liposome, and then the endocytic behavior [43][44].
An interesting delivery platform to consider is light-triggered lipid formulation. In fact, after injection of classic LNPs (locally or systemically), there is no option to physically trigger the endosomal escape. Light-triggered formulations, thanks to specific wavelengths suitable for therapeutic use, allow a precise spatiotemporal release of the cargo inside the cell, which can be needed in some diseases to specifically target organs, or simply to decrease systemic toxicity. Up to date, most of the formulations are liposomes [45], but can also be lipopolyplexes [46], and they show promising results in LNPs formulation [47]. To be triggered by light, different strategies exist based on a photosensitive molecule, e.g., verteporfin (VP), which, under a specific wavelength illumination, can generate reactive oxygen species (ROS) that destabilize the endolysosomal membrane [48][49], or with indocyanine green (ICG), which can absorb and convert light energy into heat to destabilize the membrane [50]. Moreover, lipids can also be light sensitive, e.g., allowing the surface charge of a liposome to change [51], which can serve to enhance systemic circulation when neutral, and cellular uptake/endosomal escape when charged. With a high interest and encouraging results in cancer therapy, thanks to precise drug release, light-triggered formulations have been shown to be capable of nucleic acid release, such as DNA or oligonucleotides, which can lead to a promising platform for mRNA delivery [48][50].
Nevertheless, lipoplexes mRNA therapy could still be interesting for therapeutic purposes. Recently, we showed that co-delivery with lipoplexes of non-structural protein-1 (NS1) mRNA with bone morphogenetic protein-2 (BMP-2) mRNA increases the expression of BMP-2 and osteogenic differentiation into murine pluripotent stem cells [52]. Influenza A virus (IAV) non-structural protein 1 is a multifunctional protein, helping virus replication and virulence, which is known to interact with several proteins (e.g., RIG-I) to inhibit the activation of transcription factors (IRF3/7, NF-kB, etc.) required for IFNs production, leading to anti-viral response and reduced translational activity. C2C12-BRE/LUC cells that stably express luciferase reporter gene under BMP-2 responsive elements (BRE-luciferase), were transfected with lipoplexes containing 1 µg of mRNA with a different ratio between BMP-2 and NS1. When cells are transfected with BMP-2 mRNA alone, luciferase activity decreases by 65% after 48h, and almost completely (91%) after 72 h. When transfected with 0.25 µg of NS1 mRNA and 0.75 µg of BMP2, luciferase activity was 3.5- and 4.7-fold higher at 48 h and 72 h, respectively, and the decrease of luciferase activity was lower after 48 h (54%) and 72 h (85%) (Figure 2) [53].
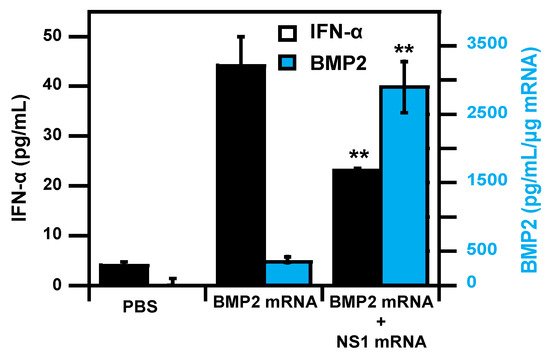
Figure 2. Co-transfection of osteoprogenitor cells with BMP2 mRNA and NS1 mRNA resulted in decreasing type I interferon response together with enhanced therapeutic BMP2 mRNA expression. Adapted from [53] with permission.
Lipopolyplexes (or lipid–polymer hybrid nanoparticles) are ternary complexes formed between a nucleic acid, a polycation, and liposomes. In 1996, Huang et al. showed that the combination of cationic polymer (poly(L-lysine), protamine) with cationic liposome resulted in smaller and more stable particles, with increased transfection efficiency in comparison to lipoplexes [54]. Since then, several studies were performed to deliver different nucleic acids with lipopolyplexes, for example, DNA [55], interfering RNAs [56][57], and mRNA [5][6][58]. Recently mRNA-loaded lipopolyplexes have emerged as a suitable therapeutic that can be used for vaccination [3].
References
- Wolff, J.A.; Malone, R.W.; Williams, P.; Chong, W.; Acsadi, G.; Jani, A.; Felgner, P.L. Direct gene transfer into mouse muscle in vivo. Science 1990, 247, 1465–1468.
- Sahin, U.; Karikó, K.; Türeci, Ö. mRNA-based therapeutics—Developing a new class of drugs. Nat. Rev. Drug Discov. 2014, 13, 759–780.
- Uchida, S.; Perche, F.; Pichon, C.; Cabral, H. Nanomedicine-Based Approaches for mRNA Delivery. Mol. Pharm. 2020, 17, 3654–3684.
- Andrée, L.; Yang, F.; Brock, R.; Leeuwenburgh, S.C. Designing biomaterials for the delivery of RNA therapeutics to stimulate bone healing. Mater. Today Bio 2021, 10, 100105.
- Perche, F.; Benvegnu, T.; Berchel, M.; Lebegue, L.; Pichon, C.; Jaffrès, P.-A.; Midoux, P. Enhancement of dendritic cells transfection in vivo and of vaccination against B16F10 melanoma with mannosylated histidylated lipopolyplexes loaded with tumor antigen messenger RNA. Nanomed. Nanotechnol. Biol. Med. 2011, 7, 445–453.
- Perche, F.; Clemençon, R.; Schulze, K.; Ebensen, T.; Guzmán, C.A.; Pichon, C. Neutral Lipopolyplexes for In Vivo Delivery of Conventional and Replicative RNA Vaccine. Mol. Ther. Nucleic Acids 2019, 17, 767–775.
- Uchida, S.; Itaka, K.; Uchida, H.; Hayakawa, K.; Ogata, T.; Ishii, T.; Fukushima, S.; Osada, K.; Kataoka, K. In Vivo Messenger RNA Introduction into the Central Nervous System Using Polyplex Nanomicelle. PLoS ONE 2013, 8, e56220.
- Lin, C.-Y.; Perche, F.; Ikegami, M.; Uchida, S.; Kataoka, K.; Itaka, K. Messenger RNA-based therapeutics for brain diseases: An animal study for augmenting clearance of beta-amyloid by intracerebral administration of neprilysin mRNA loaded in polyplex nanomicelles. J. Control. Release 2016, 235, 268–275.
- Perche, F.; Uchida, S.; Akiba, H.; Lin, C.-Y.; Ikegami, M.; Dirisala, A.; Nakashima, T.; Itaka, K.; Tsumoto, K.; Kataoka, K. Improved brain expression of anti-amyloid β scfv by complexation of mRNA including a secretion sequence with PEG-based block catiomer. Curr. Alzheimer Res. 2017, 14, 295–302.
- Kis, Z.; Kontoravdi, C.; Shattok, R.; Shah, N. Resources, Production Scales and Time Required for Producing RNA Vaccines for the Global Pandemic Demand. Vaccines 2021, 9, 3.
- Kim, J.; Eygeris, Y.; Gupta, M.; Sahay, G. Self-assembled mRNA vaccines. Adv. Drug Deliv. Rev. 2021, 170, 83–112.
- Zhang, H.; Rombouts, K.; Raes, L.; Xiong, R.; De Smedt, S.C.; Braeckmans, K.; Remaut, K. Fluorescence-Based Quantification of Messenger RNA and Plasmid DNA Decay Kinetics in Extracellular Biological Fluids and Cell Extracts. Adv. Biosyst. 2020, 4, e2000057.
- Tsui, N.B.; Ng, E.K.-O.; Lo, Y.D. Stability of Endogenous and Added RNA in Blood Specimens, Serum, and Plasma. Clin. Chem. 2002, 48, 1647–1653.
- Pardi, N.; Hogan, M.J.; Porter, F.W.; Weissman, D. mRNA vaccines—A new era in vaccinology. Nat. Rev. Drug Discov. 2018, 17, 261.
- Kowalski, P.; Rudra, A.; Miao, L.; Anderson, D.G. Delivering the Messenger: Advances in Technologies for Therapeutic mRNA Delivery. Mol. Ther. 2019, 27, 710–728.
- Midoux, P.; Pichon, C. Lipid-based mRNA vaccine delivery systems. Expert Rev. Vaccines 2014, 14, 221–234.
- Engin, A.B.; Nikitovic, D.; Neagu, M.; Henrich-Noack, P.; Docea, A.O.; Shtilman, M.I.; Golokhvast, K.; Tsatsakis, A.M. Mechanistic understanding of nanoparticles’ interactions with extracellular matrix: the cell and immune system. Part. Fibre Toxicol. 2017, 14, 22.
- Gómez-Aguado, I.; Rodríguez-Castejón, J.; Vicente-Pascual, M.; Rodríguez-Gascón, A.; Aspiazu, M.; Ángeles, S.; Del Pozo-Rodríguez, A. Nanomedicines to Deliver mRNA: State of the Art and Future Perspectives. Nanomaterials 2020, 10, 364.
- Duncan, R.; Richardson, S.C.W. Endocytosis and Intracellular Trafficking as Gateways for Nanomedicine Delivery: Opportunities and Challenges. Mol. Pharm. 2012, 9, 2380–2402.
- Varkouhi, A.K.; Scholte, M.; Storm, G.; Haisma, H.J. Endosomal escape pathways for delivery of biologicals. J. Control. Release 2011, 151, 220–228.
- Karikó, K.; Muramatsu, H.; Ludwig, J.; Weissman, D. Generating the optimal mRNA for therapy: HPLC purification eliminates immune activation and improves translation of nucleoside-modified, protein-encoding mRNA. Nucleic Acids Res. 2011, 39, e142.
- Miao, L.; Li, L.; Huang, Y.; Delcassian, D.; Chahal, J.; Han, J.; Shi, Y.; Sadtler, K.; Gao, W.; Lin, J.; et al. Delivery of mRNA vaccines with heterocyclic lipids increases anti-tumor efficacy by STING-mediated immune cell activation. Nat. Biotechnol. 2019, 37, 1174–1185.
- Verbeke, R.; Lentacker, I.; De Smedt, S.C.; Dewitte, H. The dawn of mRNA vaccines: The COVID-19 case. J. Control. Release 2021, 333, 511–520.
- Buschmann, M.; Carrasco, M.; Alishetty, S.; Paige, M.; Alameh, M.; Weissman, D. Nanomaterial Delivery Systems for mRNA Vaccines. Vaccines 2021, 9, 65.
- Evers, M.J.W.; Kulkarni, J.A.; Van Der Meel, R.; Cullis, P.R.; Vader, P.; Schiffelers, R.M. State-of-the-Art Design and Rapid-Mixing Production Techniques of Lipid Nanoparticles for Nucleic Acid Delivery. Small Methods 2018, 2, 1700375.
- Hajj, K.A.; Ball, R.L.; Deluty, S.B.; Singh, S.R.; Strelkova, D.; Knapp, C.M.; Whitehead, K.A. Branched-Tail Lipid Nanoparticles Potently Deliver mRNA In Vivo due to Enhanced Ionization at Endosomal pH. Small 2019, 15, e1805097.
- Paunovska, K.; Sago, C.D.; Monaco, C.M.; Hudson, W.; Castro, M.G.; Rudoltz, T.G.; Kalathoor, S.; Vanover, D.A.; Santangelo, P.J.; Ahmed, R.; et al. A Direct Comparison of in Vitro and in Vivo Nucleic Acid Delivery Mediated by Hundreds of Nanoparticles Reveals a Weak Correlation. Nano Lett. 2018, 18, 2148–2157.
- Tanaka, H.; Sakurai, Y.; Anindita, J.; Akita, H. Development of lipid-like materials for RNA delivery based on intracellular environment-responsive membrane destabilization and spontaneous collapse. Adv. Drug Deliv. Rev. 2020, 154–155, 210–226.
- Kirschman, J.L.; Bhosle, S.; Vanover, D.; Blanchard, E.L.; Loomis, K.H.; Zurla, C.; Murray, K.; Lam, B.C.; Santangelo, P.J. Characterizing exogenous mRNA delivery, trafficking, cytoplasmic release and RNA-protein correlations at the level of single cells. Nucleic Acids Res. 2017, 45, e113.
- Florence, A.T. Reductionism and complexity in nanoparticle-vectored drug targeting. J. Control. Release 2012, 161, 399–402.
- Torchilin, V.; Amiji, M.M. Handbook of Materials for Nanomedicine; CRC Press: Boca Raton, FL, USA, 2011; Volume 1.
- Zukancic, D.; Suys, E.J.A.; Pilkington, E.H.; Algarni, A.; Al-Wassiti, H.; Truong, N.P. The Importance of Poly(ethylene glycol) and Lipid Structure in Targeted Gene Delivery to Lymph Nodes by Lipid Nanoparticles. Pharmaceutics 2020, 12, 1068.
- Thi, T.; Suys, E.; Lee, J.; Nguyen, D.; Park, K.; Truong, N. Lipid-Based Nanoparticles in the Clinic and Clinical Trials: From Cancer Nanomedicine to COVID-19 Vaccines. Vaccines 2021, 9, 359.
- Hatakeyama, H.; Akita, H.; Harashima, H. The Polyethyleneglycol Dilemma: Advantage and Disadvantage of PEGylation of Liposomes for Systemic Genes and Nucleic Acids Delivery to Tumors. Biol. Pharm. Bull. 2013, 36, 892–899.
- Schoenmaker, L.; Witzigmann, D.; Kulkarni, J.A.; Verbeke, R.; Kersten, G.F.A.; Jiskoot, W.; Crommelin, D.J.A. mRNA-lipid nanoparticle COVID-19 vaccines: Structure and stability. Int. J. Pharm. 2021, 601, 120586.
- Perche, F.; Biswas, S.; Torchilin, V.P. Stimuli-Sensitive Polymeric Nanomedicines for Cancer Imaging and Therapy. In Handbook of Polymers for Pharmaceutical Technologies; John Wiley & Sons: Hoboken, NJ, USA, 2015; Volume 2, pp. 311–344.
- Perche, F.; Torchilin, V.P. Recent Trends in Multifunctional Liposomal Nanocarriers for Enhanced Tumor Targeting. J. Drug Deliv. 2013, 2013, 1–32.
- Brader, M.L.; Williams, S.J.; Banks, J.M.; Hui, W.H.; Zhou, Z.H.; Jin, L. Encapsulation state of messenger RNA inside lipid nanoparticles. Biophys. J. 2021.
- Torchilin, V.P. Recent advances with liposomes as pharmaceutical carriers. Nat. Rev. Drug Discov. 2005, 4, 145–160.
- Felgner, P.L.; Gadek, T.R.; Holm, M.; Roman, R.; Chan, H.W.; Wenz, M.; Northrop, J.P.; Ringold, G.M.; Danielsen, M. Lipofection: A highly efficient, lipid-mediated DNA-transfection procedure. Proc. Natl. Acad. Sci. USA 1987, 84, 7413–7417.
- Felgner, P.L.; Ringold, G.M. Cationic liposome-mediated transfection. Nat. Cell Biol. 1989, 337, 387–388.
- Rädler, J.O.; Koltover, I.; Salditt, T.; Safinya, C.R. Structure of DNA-Cationic Liposome Complexes: DNA Intercalation in Multilamellar Membranes in Distinct Interhelical Packing Regimes. Science 1997, 275, 810–814.
- Guevara, M.L.; Persano, S.; Persano, F. Lipid-based vectors for therapeutic mRNA-based anti-cancer vaccines. Curr. Pharm. Des. 2019, 25, 1443–1454.
- Wahane, A.; Waghmode, A.; Kapphahn, A.; Dhuri, K.; Gupta, A.; Bahal, R. Role of Lipid-Based and Polymer-Based Non-Viral Vectors in Nucleic Acid Delivery for Next-Generation Gene Therapy. Molecules 2020, 25, 2866.
- Franco, M.S.; Gomes, E.R.; Roque, M.C.; Oliveira, M.C. Triggered Drug Release from Liposomes: Exploiting the Outer and Inner Tumor Environment. Front. Oncol. 2021, 11, 470.
- Chen, W.; Deng, W.; Xu, X.; Zhao, X.; Vo, J.N.; Anwer, A.G.; Williams, T.; Cui, H.; Goldys, E.M. Photoresponsive endosomal escape enhances gene delivery using liposome–polycation–DNA (LPD) nanovectors. J. Mater. Chem. B 2018, 6, 5269–5281.
- Chander, N.; Morstein, J.; Bolten, J.S.; Shemet, A.; Cullis, P.R.; Trauner, D.; Witzigmann, D. Optimized Photoactivatable Lipid Nanoparticles Enable Red Light Triggered Drug Release. Small 2021, 2008198.
- Chen, W.; Deng, W.; Goldys, E.M. Light-Triggerable Liposomes for Enhanced Endolysosomal Escape and Gene Silencing in PC12 Cells. Mol. Ther. Nucleic Acids 2017, 7, 366–377.
- Aksoy, Y.A.; Yang, B.; Chen, W.; Hung, T.; Kuchel, R.P.; Zammit, N.W.; Grey, S.T.; Goldys, E.M.; Deng, W. Spatial and Temporal Control of CRISPR-Cas9-Mediated Gene Editing Delivered via a Light-Triggered Liposome System. ACS Appl. Mater. Interfaces 2020, 12, 52433–52444.
- Kontturi, L.-S.; Dikkenberg, J.V.D.; Urtti, A.; Hennink, W.E.; Mastrobattista, E. Light-Triggered Cellular Delivery of Oligonucleotides. Pharmaceutics 2019, 11, 90.
- Arias-Alpizar, G.; Kong, L.; Vlieg, R.C.; Rabe, A.; Papadopoulou, P.; Meijer, M.S.; Bonnet, S.; Vogel, S.; Van Noort, J.; Kros, A.; et al. Light-triggered switching of liposome surface charge directs delivery of membrane impermeable payloads in vivo. Nat. Commun. 2020, 11, 1–14.
- Wang, P.; Logeart-Avramoglou, D.; Petite, H.; Goncalves, C.; Midoux, P.; Perche, F.; Pichon, C. Co-delivery of NS1 and BMP2 mRNAs to murine pluripotent stem cells leads to enhanced BMP-2 expression and osteogenic differentiation. Acta Biomater. 2020, 108, 337–346.
- Wang, P.; Perche, F.; Midoux, P.; Cabral, S.; Malard, V.; Correia, I.J.; Ei-Hafci, H.; Petite, H.; Logeart-Avramoglou, D.; Pichon, C. In Vivo bone tissue induction by freeze-dried collagen-nanohydroxyapatite matrix loaded with BMP2/NS1 mRNAs lipopolyplexes. J. Control. Release 2021, 334, 188–200.
- Gao, X.; Huang, L. Potentiation of Cationic Liposome-Mediated Gene Delivery by Polycations. Biochemistry 1996, 35, 1027–1036.
- Perche, F.; Gosset, D.; Mével, M.; Miramon, M.-L.; Yaouanc, J.-J.; Pichon, C.; Benvegnu, T.; Jaffrès, P.-A.; Midoux, P. Selective gene delivery in dendritic cells with mannosylated and histidylated lipopolyplexes. J. Drug Target. 2010, 19, 315–325.
- Simion, V.; Henriet, E.; Juric, V.; Aquino, R.; Loussouarn, C.; Laurent, Y.; Martin, F.; Midoux, P.; Garcion, E.; Pichon, C.; et al. Intracellular trafficking and functional monitoring of miRNA delivery in glioblastoma using lipopolyplexes and the miRNA-ON RILES reporter system. J. Control. Release 2020, 327, 429–443.
- Gonçalves, C.; Berchel, M.; Gosselin, M.-P.; Malard, V.; Cheradame, H.; Jaffrès, P.-A.; Guégan, P.; Pichon, C.; Midoux, P. Lipopolyplexes comprising imidazole/imidazolium lipophosphoramidate, histidinylated polyethyleneimine and siRNA as efficient formulation for siRNA transfection. Int. J. Pharm. 2014, 460, 264–272.
- Van der Jeught, K.; De Koker, S.; Bialkowski, L.; Heirman, C.; Tjok Joe, P.; Perche, F.; Maenhout, S.; Bevers, S.; Broos, K.; Deswarte, K.; et al. Dendritic cell targeting mRNA lipopolyplexes combine strong antitumor T-cell immunity with improved inflammatory safety. ACS Nano 2018, 12, 9815–9829.
More
Information
Subjects:
Cell Biology
Contributor
MDPI registered users' name will be linked to their SciProfiles pages. To register with us, please refer to https://encyclopedia.pub/register
:
View Times:
1.0K
Revisions:
2 times
(View History)
Update Date:
12 Jul 2021
Table of Contents



Notice
You are not a member of the advisory board for this topic. If you want to update advisory board member profile, please contact office@encyclopedia.pub.
OK
Confirm
Only members of the Encyclopedia advisory board for this topic are allowed to note entries. Would you like to become an advisory board member of the Encyclopedia?
Yes
No
${ textCharacter }/${ maxCharacter }
Submit
Cancel
Back
Comments
${ item }
|
${ item.createdUser.fullName }
${ item.createdAt }
${ item.vote }
${ item.reply }
Delete
${ reply.createdUser.fullName }
${ reply.createdAt }
${ reply.vote }
Delete
There is no reply to this comment~
${ item.replyTextCharacter }/${ item.replyMaxCharacter }
Submit
Cancel
More
No more~
There is no comment~
${ textCharacter }/${ maxCharacter }
Submit
Cancel
${ selectedItem.replyTextCharacter }/${ selectedItem.replyMaxCharacter }
Submit
Cancel
Confirm
Are you sure to Delete?
Yes
No