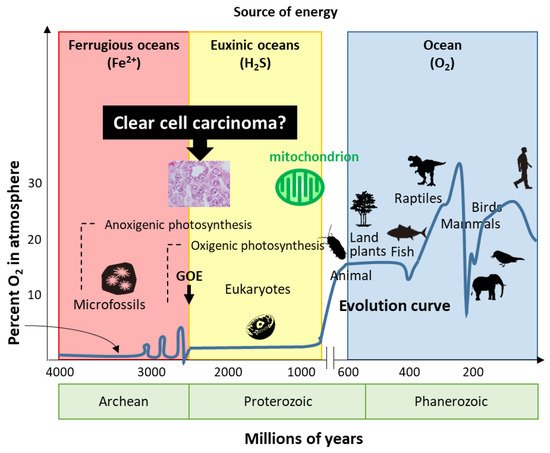
How is the expression of the ovarian CCC signature regulated? A comprehensive DNA methylation analysis showed that CCC had a profile distinct from that of other histological subtypes
[14][32]. This profile involved many genes involved in
HNF1 transcription. Both expression and DNA methylation analyses revealed that the
HNF1B signal is activated through decreased methylation in ovarian CCC. In contrast, ERa signaling is activated in endometrioid carcinoma of the ovary. In our previous report, whole exome sequencing using ovarian CCC revealed genomic abnormalities in many signals, as well as genes belonging to the
HNF1B signal in ovarian CCC
[33]. Mutations and amplification of
PIK3CA were found to occur in 54% and 26% of cases, respectively, and mutations and amplification of
KRAS were identified in 10% and 18% of ovarian CCC cases, respectively
[33]. Genomic abnormalities in
KRAS-PI3K signaling were identified in 82% of ovarian CCC cases; furthermore, 79% of CCC cases show copy number alterations in
MYC-RB signaling, including
MYC amplification and
RB1 deletion in 81% and 31% of CCC cases, respectively
[33]. The
TP53 gene was deleted, and
MDM2 and
MDM4 were amplified in ovarian CCC.
ARID1A mutations were identified in 62% of cases, and
SMARCA2 and
SMARCA4 were deleted in 21% and 41% of CCC cases, respectively
[33]; moreover, genes belonging to the SWI/SNF complex exhibited genomic abnormalities in 85% of the CCC cases. These findings suggest that the activation of
KRAS-PI3K signaling,
MYC-RB signaling, and the suppression of the
TP53 network and the SWI/SNF complex are associated with ovarian CCC
[33]. Other studies have also identified similar frequent genomic alterations in ovarian CCC. In total, 40–62% and 33–51% of CCC cases show
ARID1A and
PIK3CA mutations
[34][35].
KRAS mutations are identified in 9–17% of cases, and
TP53 mutations are observed in 5–15% of ovarian CCC cases
[34][35][36][37]. The frequency of single-nucleotide variants was significantly lower in ovarian CCC than in high-grade serous carcinoma, whereas the ratio of whole-arm copy number variants (CNVs) among all CNVs in CCC was significantly higher than that in high-grade serous carcinoma
[38]. Chromosome 20q13.2, including the
ZNF217 (Zinc finger protein 217) locus, is frequently amplified in ~36% of CCC cases
[39]. Amplifications of the
MET (chr7q31) and
AKT2 (chr19q13.2) oncogenes have been identified in 31% and 24% of ovarian CCC, respectively, and deletions of the
CDKN2A/2B (cyclin-dependent kinase inhibitor 2A/2B) tumor suppressor genes have been detected in 17% of cases
[39][40][41]. We confirmed the protein levels of activated signals in ovarian CCC. HNF1B is highly expressed, and ERa is poorly expressed in ovarian CCC
[42][43]. The expression of pAKT, which reflects
PI3K signaling, is high, and pMAPK, which reflects
KRAS signaling, is also highly expressed, similar to the genomic characteristics. Based on the immunohistochemistry of nine subunits belonging to the SWI/SNF complex, 61% of the cases exhibited the loss of at least one SWI/SNF complex subunit; thus, genomic and epigenetic abnormalities occur in ovarian CCC, which differs from the other histological types.
 +1 point
+1 point