1000/1000
Hot
Most Recent
 +1 point
+1 point
Receptors form the crux for any biochemical signaling. Receptor-like kinases (RLKs) are conserved protein kinases in eukaryotes that establish signaling circuits to transduce information from outer plant cell membrane to the nucleus of plant cells, eventually activating processes directing growth, development, stress responses, and disease resistance. Plant RLKs share considerable homology with the receptor tyrosine kinases (RTKs) of the animal system, differing at the site of phosphorylation. Typically, RLKs have a membrane-localization signal in the amino-terminal, followed by an extracellular ligand-binding domain, a solitary membrane-spanning domain, and a cytoplasmic kinase domain. The functional characterization of ligand-binding domains of the various RLKs has demonstrated their essential role in the perception of extracellular stimuli, while its cytosolic kinase domain is usually confined to the phosphorylation of their substrates to control downstream regulatory machinery. Identification of the several ligands of RLKs, as well as a few of its immediate substrates have predominantly contributed to a better understanding of the fundamental signaling mechanisms. In the model plant Arabidopsis, several studies have indicated that multiple RLKs are involved in modulating various types of physiological roles via diverse signaling routes. Here, we summarize recent advances and provide an updated overview of transmembrane RLKs in Arabidopsis.
Arabidopsis RLKs modulate growth and developmental responses by governing stem-cell maintenance, cell fate determination and patterning, male and female gametophyte development, pollen-pistil interactions, embryogenesis, hormone signaling, vascular patterning, organ development, and abscission. Some of these essential responses are discussed here.
The anther generally has four lobes and each lobe contains reproductive microsporocyte surrounded by various layers of somatic cells viz., tapetum, middle layer, endothecium, and epidermis. In Arabidopsis, multiple LRR-RLKs like excess microsporocytes1 (EMS1)/extra sporogenous cell (EXS), somatic embryogenesis receptor-like kinase 1/2 (SERK1/2), receptor-like protein kinase 2 (RPK2), barely any meristem 1/2 (BAM1/2), CLAVATA3 insensitive receptor kinase (CIK1/2/3/4), ERECTA (ER), and ERECTA-like 1/2 (ERL1/2) regulate anther development, especially, the differentiation and patterning of the somatic cell layers. EMS1/EXS was the first LRR-RLK to be identified that plays a crucial role in anther cell differentiation [1][2]. The anthers of ems1/exs mutants lack tapetum but produce large numbers of microsporocytes than the wild type. In addition, delayed expression of EMS1 in the ems1 mutant tapetal initials has been shown to aid in the generation of a functional tapetum and the diminution of microsporocyte numbers [3]. These results suggest that EMS1/EXS determines the fate of tapetal cells during early anther development. Tapetum determinant 1 (TPD1), a small secreted protein, is known to induce the phosphorylation of EMS1/EXS, thus, behaving as their ligand; and the signal is transduced downstream via phosphorylation of β-carbonic anhydrases (βCAs) [4][5]. Similarly, SERK1/2 has also been known to determine tapetal cell fate, as the anthers of serk1serk2 double mutants are phenotypically similar to that of ems1/exs mutant [6][7]. Moreover, SERK1 interacts with and transphosphorylates EMS1 to enhance its activity for guiding a co-regulatory network (Figure 1A) [8]. Corroborated by the phenotype of rpk2 mutants, it can be deduced that RPK2 is responsible for the differentiation of middle layers and tapetum during anther development. It essentially controls tapetal cell fate by triggering their degradation via modulation of the enzymes involved in cell wall metabolism and lignin biosynthesis [9] (Figure 1A). Both BAM1 and BAM2 are responsible for regulating early stages of anther differentiation, as confirmed by the lack of somatic cell layers, including endothecium, middle layer, and tapetum in bam1bam2 double mutants [10]. CLAVATA3 insensitive receptor kinases (CIK1/2/3/4) are co-receptors of BAM1/2 and RPK2, which regulate the determination of parietal cell fate and archesporial cell division [11] (Figure 1A). ERECTA (ER), ERECTA-Like 1 (ERL1), and ERL2 are also known to play essential roles in healthy anther lobe formation and anther cell differentiation via mitogen-activated protein kinases like MPK3/MPK6 (Figure 1A). The sterility of er-105 erl1-2 erl2-1 triple mutant and the phenotypic similarity of the anther lobes in single mutants of er-105 or erl1-2 or erl2-1 with that of mpk3 or mpk6 mutants suggests the correlation of these genes in the regulation of anther cell division and differentiation [12]. Further, a Lectin RLK, small, glued together, collapsed (SGC) has also been validated as a regulator of pollen development as its knockout had led to the development of small, glued-together and collapsed pollen and resulted in male sterility [13] (Figure 1A).
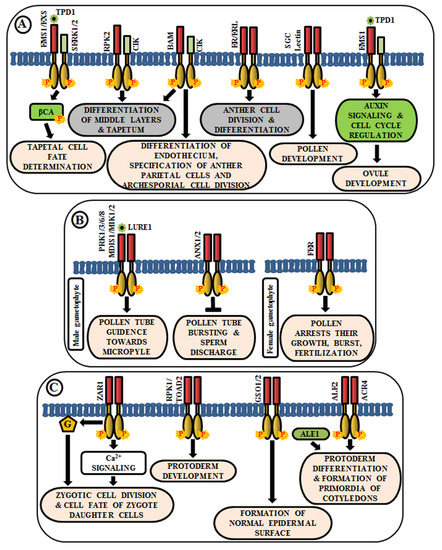
Figure 1. Arabidopsis RLKs in the regulation of growth and development. A few examples of RLKs that regulate (A) anther and ovule development, (B) pollen-pistil interaction, and (C) embryo development.
Knowledge about the role of RLKs in ovule development is very scarce. In Arabidopsis ovules, EMS1 is expressed in nucellar epidermis and chalaza, while TPD1 is weakly restricted to the distal end of integuments. Altered expression of cell-cycle genes and auxin signaling genes during ovule development, concomitant with the ectopic expression of TPD1, indicates the regulation of ovule development by TPD1-EMS1 [14] (Figure 1A).
Reproduction in angiosperms involves the release of an immobile male gamete from the pollen tube onto the compatible pistil. A fruitful pollen-pistil interaction is a pre-requisite for successful fertilization and this requires an accurate perception of ovule-emitted guidance cues by the receptors in pollen tubes. LURE1, an ovule-secreted peptide is perceived by RLKs like pollen receptor kinase 1 (PRK1), PRK3, PRK6, PRK8 in the pollen tube [15]. Recent studies ascertain the presence of other LURE receptors like Male Discoverer 1 (MDIS1), MDIS1-interacting receptor-like kinase1 (MIK1), and MIK2 [16][17]. Once the pollen tube reaches the micropyle, its growth is ceased and the sperm cells are released by its rupture. These processes are regulated by the RLK FERONIA (FER), which is expressed in the synergids of female gametophyte [18] (Figure 1B). The phenotypic study of fer mutants exhibited overgrowth of pollen tube and loss of its rupturing ability [19]. ANXUR1 and ANXUR2 (ANX1, ANX2) are homologs of FER-RLK, expressed at the tip of the pollen tube. The anx1anx2 double mutants have been found to arrest the growth of pollen tubes and promote bursting immediately after germination. These validate the clue that both FER-mediated and ANX-dependent signaling cascades act as a switch for accurate pollen tube growth and subsequent release of sperm cells for fertilization [20] (Figure 1B).
After successful fertilization, the zygote develops into embryo via repeated cell division and differentiation. Several genetic evidences suggest that multiple signaling cascades are essential for embryogenesis in Arabidopsis, and RLK is one amongst them. Predominantly, embryo development initiates from the asymmetric division of the zygote. Intriguingly, the transcript of ZYGOTIC ARREST 1 (ZAR1), a LRR-RLK, has been detected in the embryo sac before and after fertilization. It has been noticed in an eight-nucleate stage of embryo sac to different cells of mature embryo sac including the central cell, egg cell, and synergids. Even after fertilization, it was observed in the endosperm. Phenotypic analysis of zar1 mutants revealed the role of ZAR1 in the regulation of asymmetric division of zygote and determination of the cell fate of its daughter cells via the activation of calcium and G-protein signaling cascades [21] (Figure 1C). Besides ZAR1, receptor-like protein kinase 1 (RPK1) and Toadstool 2 (TOAD2) are considered indispensable for normal protoderm development, while GASSHO 1 (GSO1) and GSO2 are crucial for the formation of the proper epidermal surface during embryogenesis. The gso1gso2 double mutants have shown abnormal bending of embryos, highly permeable epidermal structure, and irregular stomatal patterning [22][23] (Figure 1C). Further, molecular analysis has detected the interaction of ALE2 (Abnormal Leaf Shape 2) and ACR4 (CRINKLY 4) with a subtilisin-like serine protease ALE1, which is essential for the formation of primordia of cotyledons during embryogenesis [24] (Figure 1C).
Coordinated cell growth, differentiationand morphogenesis are the three fundamental aspects of development that cause an organism to procure its shape and an intricate cascade of gene regulatory networks comprising RLKs are known to be implicated in this. In higher plants, all the aerial organs develop from shoot apical meristem (SAM). The maintenance of undifferentiated cells of SAM and organ formation through differentiation from the progeny cells are two processes maintained in a balance during the common developmental process. Interestingly, different RLKs are known to suffice this role. In Arabidopsis, CLAVATA1 or CLV1 (RLK), CLV2 (RLP) and CLV3 (secreted polypeptide) perform a pivotal role in meristem and organ development [25][26][27]. The CLV3 polypeptide acts as a ligand for CLV1 and CLV2 complex. This ligand-receptor binding promotes the activation of cytosolic kinase domain of CLV1 and subsequently, it initiates a signal transduction cascade to control gene expression and stem cell fate in the SAM by elevation of cytosolic calcium as secondary messengers [25][28][29] (Figure 2A). Meristematic receptor-like kinase (MRLK), a LRR-RLK expressed in shoot and root apical meristems, interacts with and phosphorylates a MADS-box transcription factor, AGL24, to regulate floral transition [30] (Figure 2A). Another LRR-RLK, ERECTA, which is expressed in the entire shoot apical meristem and developing organs, monitors organ shape and inflorescence architecture, upon the perception of epidermal patterning factors (EPFs)/EPF-like proteins (EPFLs) [31] (Figure 2A). Moreover, mutants of ERECTA-family LRR-RLKs conferred extreme dwarfism and abnormal flower development, suggesting that ERECTA-family RLKs control cell proliferation as well as organ growth and patterning like stomata formation, the shoot apical meristem (SAM) and flower development [32]. ERECTA can form complexes with a range of co-receptors like SERKs and transmembrane receptor-like proteins like Too Many Mouths (TMM) to activate the signaling pathway [33][34]. Botrytis-induced kinase 1 or BIK1, an RLCK, interacts and phosphorylates ER-family proteins to modulate leaf morphogenesis and inflorescence architecture [35] (Figure 2A).
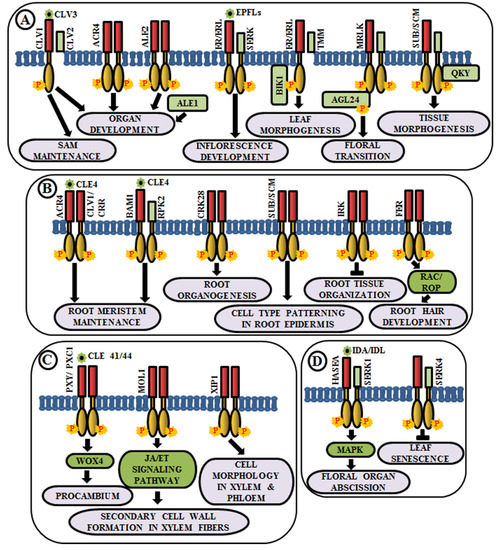
Figure 2. Arabidopsis RLKs in the regulation of growth and development. A few examples of RLKs that regulate (A) shoot development, (B) root development, (C) vascular tissue development, and (D) organ abscission.
Similar to aboveground organ development, several studies demonstrated the utmost importance of multiple RLKs in root development. Arabidopsis CRINKLY 4 (ACR4) is involved in the formation of proper lateral roots and columella stem cell differentiation in the root apical meristem [36][37]. ACR4 can regulate root meristem maintenance in response to the CLE4 peptide by forming heterodimers with CLV1 [38] (Figure 2B). Besides, ACR4, abnormal leaf shape 1 (ALE1) (a member of subtilisin-like serine protease family), and ALE2 (RLK) have been known to share partial overlapping roles in the formation of leafy organs [24] (Figure 2A). Similar to ACR4, cysteine-rich receptor-like kinases (CRKs), a member of one of the largest RLK families, is involved in root organogenesis. The crk28 mutants have displayed longer and branched roots, while CRK28 overexpression lines have shown the contrasting phenotype, i.e., delayed root growth and reduced lateral root formation [39] (Figure 2B).
Plasmodesmata are microchannels between two cells, through which trafficking of molecules occur. STRUBBELIG (SUB) is a RLK involved in inter-cell layer signaling which is required for tissue morphogenesis. The sub mutants have shown defects in floral organ shape, integument initiation, and outgrowth, asymmetry in leaf shape and stem morphology, as well as a reduction in plant height. This indicates the functional role of SUB across several cells in the floral meristem, ovule, and shoot apex [40][41]. Further genetic screening has led to the identification of a putative membrane-anchored C2-domain protein, encoded by QUIRKY (QKY), which is known to act as a downstream component of SUB signaling [42]. SUB and QKY interact in plasmodesmata to promote tissue morphogenesis (Figure 2A). Apart from aerial organs, SUB or SCRAMBLED (SCM) also regulates cell-type patterning in the root epidermis [43] (Figure 2B). The BAM1 (barely any meristem 1), a member of CLV1 class LRR-RLKs, is expressed preferentially in the quiescent center and its surrounding stem cells at the root tip and known to bind to the CLE peptide. BAM1 is capable of forming heteromeric complexes with RPK2 and inhibit cell proliferation in the root meristem [44] (Figure 2B). Inflorescence and root apices receptor kinase (IRK), a typical meristematic LRR-RLK, is known to be expressed in the outer plasma membrane of root endodermal cells and negatively regulates cell division to maintain tissue organization [45] (Figure 2B). Further, FERONIA (FER) receptor-like kinase functions upstream of Rho-like small G-protein or RAC/ROP during reactive oxygen species (ROS)-mediated root hair development. The FER activates RAC/ROP by GDP-GTP exchange to stimulate NADPH oxidase for ROS formation [46] (Figure 2B).
The development of xylem and phloem from the vascular meristem is a multifaceted process. The RLK, phloem intercalated with xylem (PXY), maintains cell polarity during vascular development, which is ascertained by the presence of partially interspersed xylem and phloem, and irregular vascular development in pxy mutants [47]. The ligand for PXY receptor is tracheary element differentiation factor (TDIF), a peptide, which is encoded by CLAVATA3/ESR 41/44 (CLE41/44) genes [48]. The PXY-TDIF interaction activates the WUSCHEL-related homeobox 4 (WOX4) signaling pathway to regulate cell division in the procambium. Another LRR-RLK, PXY/TDR-CORRELATED (PXC1), acts as a positive regulator of secondary cell wall formation in xylem fibers [49] (Figure 2C). The CLE41/PXY/WOX4 cascade is antagonistically directed by the LRR-RLK more lateral growth 1 (MOL1), via regulating the stem cell homeostasis within the cambium. This MOL1 also attenuates ethylene and jasmonic acid hormone signaling pathways that positively influence cambium activity [50] (Figure 2C). The maintenance of the cell morphology organization during vascular development is accomplished by a RLK, xylem intermixed with phloem 1 (XIP1). Genetic evidences also unveil that XIP1 prevents ectopic lignification in phloem cells [51] (Figure 2C).
Arabidopsis LRR–RLK HAESA (formerly named RLK5) exhibits developmentally regulated expression in the abscission layers of floral organs. The antisense suppression of the HAESA is known to delay the abscission of floral organs such as sepals, petals, and stamens [52]. Inflorescence deficient in abscission (IDA) and IDA-Like (IDL) proteins are considered as the ligands of HAESA (HAE) and HAESA-Like RLKs [53] (Figure 2D). The phenotypic analysis of ida mutant and overexpression of IDA gene validates the role of HAE in floral organ abscission via IDA/IDL perception. A phosphorylation-based activation mechanism of HAE leads to the stimulation of a MAP kinase-signaling cascade and initiates cell wall hydrolysis at the base of the abscising organs. SERK1 acts as a co-receptor of HAE and allows the binding of IDA, eventually leading to floral abscission pathway [54][55]. In contrast, an early leaf senescence phenotype observed in serk4-1 knockout mutant indicates that SERK4 acts as a co-receptor in negatively regulating leaf senescence, as well [56] (Figure 2D).
Brassinosteroids (BRs) are essential polyhydroxylated steroidal phytohormones crucial for plant development. The developmental defects of BR biosynthetic and signaling mutants are mostly similar, which include dwarfism, severely stunted and rounded leaf with a shorter petiole, delayed flowering, photomorphogenic malfunctions as well as senescence and reduced male fertility. The first BR signaling gene, whose mutation showed these phenotypes, has been named as brassinosteroid insensitive 1 (BRI1) [133]. BAK1 (BRI1-associated receptor kinase 1), a co-receptor of BRI1, is involved in BR perception and signaling via heterodimerization with BRI1 [57][58]. In addition, a close homologue of BRI1, BRI1-like receptor kinase (BRL1) is also responsible for BR perception [59] (Figure 3A). BAK1-associating receptor-like kinase 1 (BARK1), a LRR-RLK, specifically binds to BAK1 and its homologs. Overexpression of BARK1 enhances primary root growth and these roots are hypersensitive to BR-induced root growth inhibition, suggesting the role of BARK1 in BR-mediated lateral root development via auxin signaling [60] (Figure 3A). Apart from these, evidence achieved from bir1 mutants helps us to comprehend how it modulates immune response pathways and plant architecture as an interacting partner of BAK1 [61]. A member of somatic embryogenesis receptor, SERK3 acts as a co-receptor, which directly interacts with BRI1 [62] (Figure 3A).
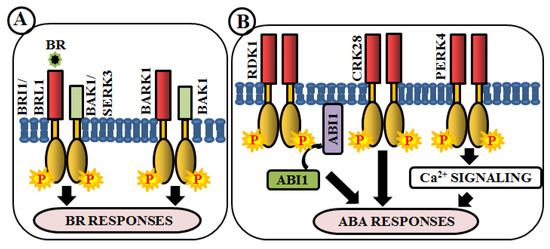
Figure 3. Arabidopsis RLKs in brassinosteroid (BR) and abscisic acid (ABA) signaling. RLK-mediated phosphorylation-based signaling circuits regulate BR (A) and ABA (B) responses.
Abscisic acid (ABA) is yet another vital phytohormone involved in the regulation of plant abiotic stress-related phenotype as well as developmental processes. Unlike BR, in Arabidopsis, RLKs are not accountable for direct ABA perception. A LRR-RLK, receptor dead kinase 1 (RDK1) is involved in ABA signal transduction via interaction with abscisic acid insensitive 1 (ABI1), a type 2C protein phosphatase, in the plasma membrane. Predominantly, this interaction is enhanced by exogenous application of ABA, underpinning the involvement of RDK1 to recruit ABI1 to the plasma membrane [63] (Figure 3B). Most recently, a cysteine-rich receptor-like kinase, CRK28, has shown an indirect relationship with ABA. The CRK28 overexpression lines have displayed slow root growth, reduced lateral root formation, and also ABA hypersensitivity; thereby being an important modulator of ABA signaling [39] (Figure 3B). PERK4 is also known to play an important role in ABA response. The perk4 mutants have shown reduced sensitivity to ABA concerning seed germination, seedling growth, and primary root tip growth. Moreover, perk4 mutant cells have retained lower cytosolic calcium concentration and Ca2+ channel currents. These results suggest that PERK4 contributes to the early stage of ABA signaling and inhibits root cell elongation via intracellular calcium signaling [64] (Figure 3B). Other RLKs like CRK5, CRK36, LRK10L1.2, and RPK1 are also known to be involved in ABA signaling during response to drought and oxidative stresses.





