Cancer is a condition where cells proliferate uncontrollably [
1,
2]. It is the leading cause of death in economically developed countries and the second leading cause of death in developing countries [
3]. Colorectal cancer (CRC) is the third most common cause of cancer death globally, with an estimated 2.2 million new cases and 1.1 million deaths expected over the next decade [
4]. CRC accounts for 9.2% of cancer-related deaths, making it the second leading cause of cancer death [
5,
6,
7]. Both genetic and environmental factors play a role in CRC causation [
8]. Chronic colitis due to inflammatory bowel disease (IBD) is also associated with an increased risk of CRC [
9]. CRC has a poor prognosis, and there is a critical need for new diagnostic and prognostic biomarkers to reduce CRC-related deaths [
10,
11]. Cytokine networks are largely involved in the prognosis and pathogenesis of CRC [
12]. Tumors express cytokines with both antitumor and pro-tumor properties [
13]. Cytokines’ ability to stimulate oncogenic signaling has shifted research focus towards their role in promoting cell proliferation and survival during tumorigenesis [
14]. Immune cells activated in response to the tumor produce and release growth factors and cytokines to regulate the inflammatory environment in tumor tissues [
15]. These inflammatory cytokines direct DNA damage in the epithelium [
16]. They imply the pathogenesis and prognosis of CRC [
12]. Interleukins have distinct roles in CRC progression through tumorigenesis [
14] and are also involved in tumor growth, cancer cell invasion, and metastasis, while also inhibiting cancer through complex pathways [
15]. In recent years, interleukins have gained significant attention due to their distinct roles in providing a new and promising strategy for CRC treatment [
17].
2. Molecular Pathways and Cytokine Role in CRC
The tumor microenvironment (TME) plays an important role in the initiation and growth of tumors into metastatic states [
35]. Tumor cells secrete cytokines that stimulate self-proliferation, drug resistance, and activation and differentiation of other cells in the tumor microenvironment [
36,
37]. Neutrophils are triggered by complement immunity and release IL-1β, which can activate myeloid cells to produce IL-6 and increase the IL-17A response. IL-33 can be secreted by vascular endothelial cells and tumor cells [
17]. The host’s immune response causes chronic inflammation from time to time, which leads to tumor growth primarily due to interactions between tumors, immune, and other cells moderated by cytokine [
38,
39]. Cytokines alert immune cells in the presence of tissue damage and infections to stimulate the cells to release more cytokine signals leading to chronic inflammation [
38]. Chronic inflammation promotes diverse cytokine secretion during various stages and types of cancer [
32]. It causes DNA damage and DNA mutation by reactive oxygen and nitrogen species, along with the alteration of the epigenome of the cells [
40,
41].
Immunological biomarkers can be predictive, prognostic, or both [
42]. Biomarkers are genes, gene products, cells, enzymes, molecules, or hormones that can be detected in tissues and body fluids (blood, urine, etc.) [
43]. Cancer biomarkers are usually produced by the tumor or other body cells in response to the tumor [
44]. They can be used for early cancer detection and patient state prediction [
45]. Certain biomarkers can reveal genetic changes in cancerous cells, such as gene rearrangements or amplifications, mutations, and cell division, enabling cancer detection and response prediction to various treatments [
46,
47]. Prognostic immune cells include tumor-infiltrating lymphocytes, eosinophils, neutrophilic granulocytes, macrophages, and dendritic cells [
48,
49,
50,
51]. The epithelium of a tumor activates stromal cells to release cytokines, such as transforming growth factor, and cell signaling factors to develop a microenvironment for tumor progression and metastasis [
52,
53,
54].
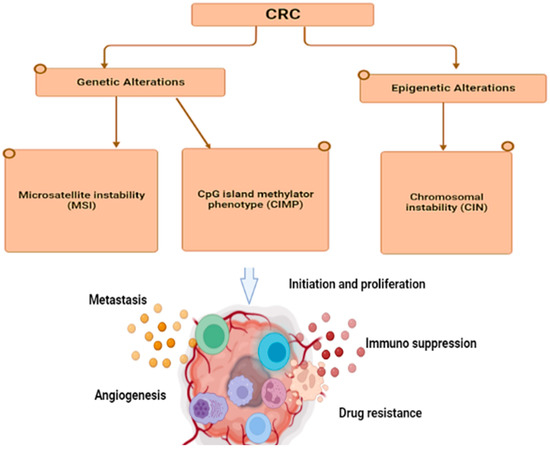
The development of colorectal cancer is also caused by specific mutations in oncogenes, tumor-suppressor genes, and genes associated with DNA repair mechanisms. However, approximately 70% of colorectal cancer cases follow a specific mutation and are transformed into a specific morphological sequence that starts with polyps and abnormal crypts, which then develop into early adenomas. The adenoma then progresses to advanced adenomas, eventually leading to colorectal cancer [
55]. Therefore, genome instability is a fundamental feature of CRC. The pathogenic mechanisms leading to this situation can be divided into three different pathways (
Figure 1): chromosomal instability (CIN), microsatellite instability (MSI), and CpG island methylator phenotype (CIMP) [
56].
Figure 1. Molecular alterations and Genetic changes cause various pathways: chromosomal instability (CIN), microsatellite instability (MSI), and CpG island methylator phenotype (CIMP,) which lead to metastasis, angiogenesis, drug resistance, immunosuppression, and inflammation in tumors.
Mesenchymal cells of the gut, including endothelial cells, smooth muscle cells, mucosal immune cells, and subepithelial myofibroblasts, are combined with the tumor epithelial cells to regulate TME for the progression of CRC [
57,
58].
Table 1 represents main cancer cell types with the signaling pathways and target cells.
Table 1. Cancer cell types in TME and the specific soluble factors released by them. Soluble factors involved in the promotion of cancer stem ness pathways.
Many polymorphisms and somatically altered genes in CRC affect the KRAS- (kirsten rat sarcoma viral oncogene homolog), MYC- (cellular myelocytomatosis oncogene), Wnt-, mitogen-activated protein kinase (MAPK)-, or TGF-β/bone morphogenetic protein (BMP)-signaling pathways. For example, the nuclear factor κB (NF-κB), JNK, AP-1, and p38 MAPK pathways are activated by IL-1 receptor cytokine in CRC along with the activation of the Wnt signaling pathway by phosphorylation of GSK3β [
12].
3. Advancements in Cytokine Detection and Monitoring Clinically
The study of cytokine secretion can lead to improved understanding of disease mechanisms and the development of new treatments. Elevated cytokine levels are a hallmark of many diseases such as diabetes, sepsis, cardiovascular diseases, neurodegenerative diseases, and cancers, and can be used as potential biomarkers [
64,
65,
66,
67,
68]. A study found that a combination of IFN-γ, IP-10, ferritin, and 25-hydroxyvitamin D can be used to diagnose pediatric tuberculosis and to differentiate between TB and latent TB infection [
69]. Inflammatory cytokines such as IL-6, IL-10, IL-1, TNF-α, and TGF-β are linked to the progression of heart failure and chronic kidney dysfunction, which are major global health problems with significant economic impacts [
70]. Furthermore, inflammatory bowel diseases such as ulcerative colitis and Crohn’s disease result from an imbalance in pro- and anti-inflammatory cytokine interactions [
29]. However, excessive cytokine release into the bloodstream from immune cells can cause cytokine release syndrome during immunotherapy and infections [
71,
72]. This makes detecting cytokines challenging due to their low concentration, instability, and complex networks [
73]. Cytokine imbalances can also cause illnesses and prompt the need for precise and early treatment in severe conditions, so there is a demand for accurate, fast, and sensitive cytokine screening methods [
74,
75].
Common methods for cytokine quantification in clinical practice include ELISA and ELISpot [
76]. Due to their crucial role in disease and human health, researchers are striving to develop tools for sensitive, multiplex cytokine detection [
77]. For instance, recent studies have explored deployable devices with immunosensors on fiber optics [
78] and stainless steel [
79] for cytokine monitoring in mouse brain and spinal cords at pg/mL levels. An impedance aptasensor was developed for a highly sensitive detection of IL-6, with a detection limit of 1.6 pg/mL and a linear response of 5 pg/mL to 100 ng/mL [
80]. A microfluidic technology based on single plex was designed to eliminate cross-reactivity and detect chemokines and cytokines in human and mouse samples [
81]. Another chemiluminescent nanozyme immunoassay was developed for simultaneous detection of two chicken cytokines (IL-4 and IFN-γ) in serum samples with a range of 0.01–60 ng/mL for IFN-γ and IL-4, respectively, and detection limits of 2.9 pg/mL for IFN-γ and 3.2 pg/mL for IL-4 [
82]. Clinical detection of serum biomarkers such as CA 19–9 and CA724 has a low specificity and sensitivity in gastric cancer diagnosis [
83]. Studies show that the sensitivity, accuracy, and specificity of interleukins are as good as that of other markers such as carcinoembryonic antigen (CEA) and CA 19–9 [
84]. This suggests that IL-6 may be a reliable biomarker for gastric cancer detection and diagnosis.
IL detection is commonly used as a biomarker for infections in clinical applications [
85,
86]. The most widely used tumor marker is a combination of ILs and CEA [
87]. Tumor markers, found in body fluids, are widely used for early diagnosis, prognostic evaluation, and treatment monitoring of tumors [
88,
89], but CEA lacks specificity [
90]. Combining inflammatory factors and tumor markers has shown a high potential in diagnosing pancreatic and CRC [
91,
92]. IL detection has also been analyzed for its individual role in the diagnosis of lung cancer and predicting lymph node metastasis [
93]. They have shown promising results as a cancer biomarker in various types of cancer including gastric cancer, pancreatic cancer, and CRC. The use of ILs combined with other tumor markers such as CEA has the potential to enhance the accuracy of cancer diagnosis. In the case of thyroid cancer, the combination of IL-13 and IL-8 has been found to be effective in identifying the disease [
94]. However, further research is needed to fully understand the genetic basis of interleukin levels as a cancer risk factor and to account for other factors that can impact the levels of interleukins in the body [
95].
Alternation in the expression of interleukins such as IL-17, IL-22, and IL-1ß has been associated with various types of cancer [
96,
97]. Interleukins regulate the tumor microenvironment and are involved in tumor development and progression [
98,
99]. Not only locally, but their actions also occur distantly through circulation [
100]. Interleukins promote cancer development by counteracting the immune response, mobilizing stromal, and immunosuppressive cells that support the tumor, inducing angiogenesis, and altering the response to therapeutic agents [
101,
102]. Their production is also triggered due to changes induced by cancer, alterations in metabolism, cell death, oxygen deficiency, and usage of anticancer drugs [
102].
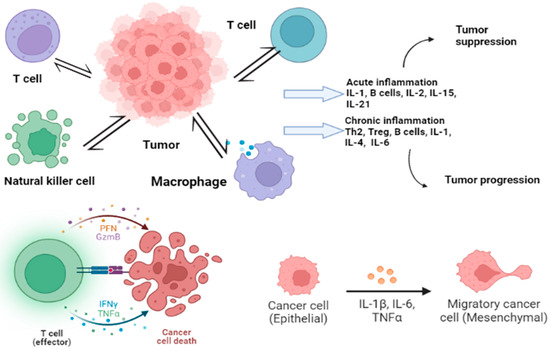
Figure 2 shows the immune response of cancer with cytokine release. The control of widespread cancer includes effective tools for the betterment of cancer burden by knowledge, early detection, suitable therapy along with a regular follow-up, and forecast measures by using cancer biomarkers [
103]. The analysis of cytokines along with cancer-specific biomarkers has been put forward to improve cancer detection [
32].
Figure 2. Immune cells influencing the tumor, leading to acute and chronic inflammation. Many cytokines (shown in form of colored dots in the figure) play their roles in this regard to either suppress or progress the tumor.
4. Cytokines’ Role in CRC
Some cytokines (other than interleukins) are also associated with the immune regulation of tumor cells and are highly expressed as compared to normal cells. They include forkhead box P3 (FOX P3), tumor necrosis factor-α (TNF-α), and interferon-gamma (IFN-γ).
4.1. Forkhead Box P3 (FOXP3)
The FoxP3 gene is expressed in regulatory T cells and is associated with cell development, transcription regulation, and DNA repair [
104,
105,
106]. In addition to its basic role in immune responses, FOXP3 also plays a significant role in cancer development [
107]. A high level of FOXP3 expression was observed in tumor cells compared to tumor-surrounding tissues, as detected through an immunohistochemistry assay [
108]. FOXP3 levels are also higher in colorectal cancer tissues than in normal colorectal tissues [
109], and its expression is associated with a poor prognosis compared to patients with low FOXP3 expression [
110]. However, a high level of FOXP3 in tumor cells is associated with longer and disease-free survival [
108]. Intra-tumoral CD4+ and FOXP3+ cell infiltration can be the most meaningful predictive factor in CRC patients [
111]. Studies show that FOXP3 expression by cancer cells results in the secretion of cytokines such as IL-10 and TGFβ into the tumor microenvironment, which suppresses immunity [
112].
4.2. Tumor Necrosis Factor-α (TNF-α)
Tumor necrosis factor-α (TNF-α) is a cytokine produced by macrophages and involved in various immune regulations [
113]. It promotes the process of epithelial-to-mesenchymal transition (EMT) in colorectal cancer, thus promoting the metastasis of colorectal cancer [
114,
115,
116]. Single-nucleotide polymorphisms in TNF-α genes are reportedly associated with the prognosis, therapy response, and survival of cancer patients [
117]. High TNF-α gene expression is associated with Stage III and IV neoplasms compared to earlier tumor stages, and TNF-α expression is increased in the serum of CRC patients [
118,
119]. Statistical analysis shows that TNF-α mRNA expression levels are significantly higher in CRC compared to normal CRC tissue, and CRC patients with low TNF-α serum levels have a significantly higher survival rate compared to patients with high levels of TNF-α [
118,
120]. The examination of TNF-α levels in plasma can be used as a diagnostic factor for CRC instead of using other invasive tests [
121].
4.3. Interferon-Gamma (IFN-γ)
The genetic variations in interferon-gamma (IFN-γ) and its receptor (IFN-γR) subunits are strongly associated with the risk of colorectal cancer and patient survival after diagnosis [
122]. IFN-γ is a major activator of macrophages and an inducer of Class II major histocompatibility complex (MHC) molecules [
123], with immune regulatory, antiviral, and antitumor properties [
124,
125]. Studies have shown that the deficiency of IFN-γ or its receptor promotes the development of colorectal cancer, whereas its specific expression activates innate immunity and inhibits tumorigenesis [
122,
124]. The specific expression of IFN-γ activates innate immunity and inhibits tumorigenesis [
124] However, IFN-γ signaling can also compromise antitumor immunity by inducing immune checkpoint inhibitory molecules on T and tumor cells [
125]. Furthermore, IFN-γ acts as a cytotoxic cytokine and initiates apoptosis in tumor cells [
126]. The
IFN-γ/
Janus Kinase 2 (JAK)/
signal transducer and activators of transcription (STAT) signaling pathway has been shown to induce programmed death-ligand 1 (PD-L1) expression in myeloid leukemia cells, pancreatic, and gastric cancer [
127,
128]. Increased PD-L1 expression levels have been found to be associated with a poor prognosis in patients with CRC [
129]. Previous studies have also reported that PD-L1 expression on tumor-infiltrating immune cells is correlated with the survival of patients with CRC [
130].
5. Interleukins in Colorectal Cancer
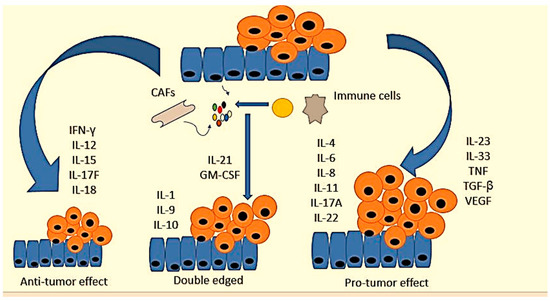
There are various interleukin families involved in CRC progression that have been studied as biomarkers (
Table 2). They play different roles in colorectal cancer. For example, IFN-γ, interleukin-12 (IL-12), IL-15, IL-17F, and IL-18 inhibit CRC development [
131,
132,
133,
134]. On the other hand, IL-4, IL-6, IL-8, IL-11, IL-17A, IL-22, IL-23, IL-33, TNF, TGF-β, and
vascular endothelial growth factor (VEGF) are pro-tumorigenic genes [
135,
136]. The contribution of IL-1, IL-9, IL-10, IL-21, and
granulocyte-macrophage colony-stimulating factor and sargramostim (GM-CSF) to intestinal cancer remains unclear [
135].
Figure 3 shows certain cytokines and their role in CRC.
Figure 3. Cytokine networks in the pathogenesis of colorectal cancer. Cytokines expressed by tumor and/or stromal cells cluster to form networks with antitumor, pro-tumor, or bivalent properties. IFN-γ, interleukin-12 (IL-12), IL-15, IL-17F, and IL-18 inhibit CRC development. IL-4, IL-6, IL-8, IL-11, IL-17A, IL-22, IL-23, IL-33, TNF, TGF-β, and VEGF are pro-tumorigenic. The contribution of IL-1, IL-9 IL-10, IL-21, and GM-CSF to intestinal cancer remains unclear.
Table 2. Major interleukins involved in CRC progression and studied as biomarkers.
|
Cytokine
|
Functional Effect in CRC
|
Expression Patterns
|
Reference
|
|
IL-1α
|
Promotes metastasis and the chemosensitivity
|

|
[136,137]
|
|
IL-1β
|
Promotes the proliferation of colon cancer cells, tumorigenesis, and alters the tumor microenvironment
|

|
[137,138,139]
|
|
IL-18
|
Antitumorigenic properties and release of other signals
|

|
[140,141]
|
|
IL-2, IL-7, IL-9, IL-15
|
Antitumor activity, promote EMT, proliferation, invasion, and metastasis
|
IL-4, IL-7 upregulated, IL-9 downregulated, IL-2 in between
|
[142,143,144,145,146,147]
|
|
IL-21
|
Activation of immune response biomarkers
|

(Potential for biomarker)
|
[148,149,150]
|
|
IL-6
|
Promotes mitosis, proliferation, metastasis, migration, and angiogenesis
|
|
[151,152,153,154]
|
|
IL-11
|
Facilitates the proliferation of CRC
|

|
[155,156,157]
|
|
IL-8
|
Promotes cell proliferation, angiogenesis, cancer metastasis, chemoresistance, antianoikis, maintains CCSC properties
|

|
[158,159,160]
|
|
IL-10
|
Pathogenesis and progression
|

|
[141,161,162,163]
|
|
IL-22
|
Dominant role in CRC tumorigenesis, antiapoptosis, and cell proliferation
|

|
[164,165,166]
|
|
IL-17a
|
Promotes cell cycle progression and angiogenesis
|

|
[167]
|
|
IL-17b
|
Promotes tumor
|

|
[168,169,170,171]
|
|
IL-4
|
Overexpressed in early CRC, tumor development
|

|
[172]
|
|
IL-23
|
Overexpressed in CRC tissue and predictive for CRC metastasis
|

|
[173,174,175,176]
|
Upwards arrow just showed upregulation of the genes while downwards show suppression or downregulation.