Anaphylaxis is one of the most life-threatening and intensive allergic reactions. Unlike anaphylactoid reaction, it is an immunoglobulin E-mediated response. Its symptoms can occur in multiple organ systems, such as cutaneous, respiratory, cardiovascular, and others. Mast cells together with basophils are the first cells that are responding to IgE-mediated anaphylaxis. Mast cells (MCs) are the immune cells distributed throughout nearly all tissues, mainly in the skin, near blood vessels and lymph vessels, nerves, lungs, and the intestines. Although MCs are essential to the healthy immune response, their overactivity and pathological states can lead to numerous health hazards. The side effect of mast cell activity is usually caused by degranulation. It can be triggered by immunological factors, such as immunoglobulins, lymphocytes, or antigen–antibody complexes, and non-immune factors, such as radiation and pathogens. An intensive reaction of mast cells can even lead to anaphylaxis, one of the most life-threatening allergic reactions. What is more, mast cells play a role in the tumor microenvironment by modulating various events of tumor biology, such as cell proliferation and survival, angiogenesis, invasiveness, and metastasis.
1. Mast Cells as a Therapeutic Target in Allergic Inflammation
Mast cells exhibit a great potential to produce mediators playing a key role in allergic-related inflammatory diseases. Among them, we differentiate biogenic amines, proteoglycans, lysosomal enzymes, lipid mediators, cytokines, growth factors, mitogens, chemokines, etc. [
42]. Allergic diseases, such as asthma, allergic rhinitis, atopic dermatitis, and others, despite occurring in different organs, share similar mechanisms.
Exposure to the factors, such as food, drugs, or plants, causes a sequence of events resulting in severe symptoms. However, according to clinical studies, in most cases the cause of anaphylaxis is undetermined (idiopathic anaphylaxis) [
44]. The general mechanism of this process starts with allergen-specific IgE antibodies binding to high-affinity Fc receptors (FcεR1) present on the MCs surface. This leads to activation and degranulation in connective or mucosal tissues in which they reside. During degranulation, MCs release mediators such as histamine, leukotrienes, prostaglandins, cytokines, proteases, kinases, and nitric oxide. After some time, other cells of the immune system join the response stimulated by mediators. In this group, special attention should be paid to eosinophils participating in allergic inflammation [
45]. Their reaction is induced by allergen-specific T helper 2 cells and Th2 cytokines [
46]. Eosinophil response to inflammation state can lead to eosinophilia, intensifying the inflammation even more, and thus increasing the intensity of the symptoms of anaphylaxis.
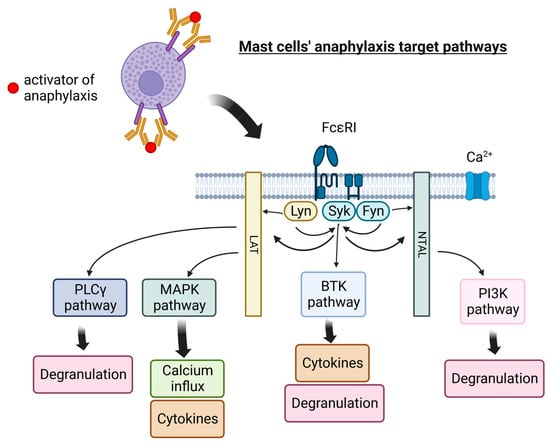
Currently, therapeutic options for the treatment of idiopathic anaphylaxis are limited with variable efficacy. Popular targets for anaphylaxis management are shown in Figure 1.
Figure 1. Mast cells’ pathways targeted in anaphylaxis therapy. PLCγ—Phosphoinositide-specific phospholipase C, MAPK—mitogen-activated protein kinases, BTK—Bruton’s tyrosine kinase, LAT—linker for activation of T cell, NTAL—non-T cell activation linker, PI3K—phosphoinositide 3-kinase, Lyn—Lyn kinase (Src family), Syk—Syk tyrosine kinase, Fyn—Fyn tyrosine kinase, FcεRI—a high-affinity IgE receptor. Figure created with BioRender.com.
Epinephrine is the first-choice medicine for anaphylaxis [
47]. It prevents hypotension and laryngeal edema, the life-threatening symptoms of the reaction. Epinephrine is one of the adrenergic drugs acting on several receptors. Its action on the α1 receptor increases vasoconstriction and peripheral vascular resistance while reducing mucosal edema. The effect on the α2 receptor reduces the release of insulin and norepinephrine. Epinephrine also acts on β1 receptors to increase inotropy and chronotropy, and β2 receptors to reduce mediator release and increase bronchodilation, vasolidation, and glyconeolysis [
48]. Although epinephrine has many advantages, some patients are showing symptoms refractory to epinephrine, such as bronchospasm. In those cases, 2-agonists, e.g., albuterol, can be used [
49]. Patients with respiratory symptoms can also benefit from high-flow oxygen. The epinephrine injection is often followed by further symptom management. At this point, corticosteroids and antihistamines apply.
Monoclonal antibodies are a novel approach in the treatment of anaphylaxis symptoms. In the group consisting of omalizumab, benralizumab, reslizumab, mepolizumab, and dupilumab, only dupilumab showed a negative signal for anaphylaxis [
50]. Dupilumab is a humanized IgG monoclonal antibody that binds to IL-4Rα, blocking IL-4 and IL-13 intracellular signaling. In the newest case report, dupilumab was effective in preventing recurrent anaphylaxis and in treating severe asthma [
51].
Inhibiting FcεRI-mediated signaling is one of the approaches that could be efficient in preventing anaphylaxis. The studies show that Sirtuin 6 can act as a negative regulator in this signaling pathway; therefore, it can be considered as a new therapeutic strategy for anaphylaxis [
52]. There exists a considerable body of literature on Bruton’s tyrosine kinase (BTK), which is an enzyme essential for high-affinity IgE receptor (FcεRI) signaling. To address BTK inhibitors’ efficacy against anaphylaxis, several FDA-approved substances were tested. Ibrutinib (Ibrivuca) is a drug that is FDA-approved for the treatment of mantle cell lymphoma, chronic lymphotic leukemia, and Waldenstroms macroglobulinemia. In the clinical studies completed in 2018 (NCT03149315), it was tested in 2 standard doses to check if it can limit allergic reactions. The studies showed reduction/elimination of skin-prick-test activity to allergens in healthy allergic adults. Acalabrutinib (Calquence) (NCT05038904) and Tirabrutinib (Velexbru) (NCT04947319) are other drugs from the group of BTK inhibitors currently tested to prevent anaphylaxis, approved or in the process of FDA approval for tumor treatment. The method introduced by Dispenza et al. has the advantage that they used an injection of human CD34+ cells into NSG-SGM3 mice, thus obtaining a model for testing the inhibition of anaphylaxis by BTK inhibitor drugs [
53]. Studies on mice confirm the effectiveness of these drugs in a new application and are the basis for further research. In the research from 2022 [
54], the authors suggested that BTK inhibitors decrease mast cell degranulation and hypothermia, but not in all of the conditions applied. In the same paper, they implied that a combination of histamine receptor 1 antagonists, β-adrenergic agonists, and a spleen tyrosine kinase (Syk) inhibitor could completely inhibit IgE-mediated hypothermia.
Several natural active compounds show great potential in managing anaphylaxis symptoms. Anemoside B4 is a triterpenoid abundant in the roots of
Pulsatilla chinensis [
55]. In both in vitro and in vivo studies, it was noted that Anemoside B4 can limit (IgE)-mediated allergic responses by inhibiting cell degranulation, calcium influx, and PLC/IP3 and JAK/STAT3 phosphorylation. Fucoxanthin is another natural compound with promising anti-anaphylactic properties [
56,
57]. The studies show that the use of fucoxanthin allows the management of cytokine production, the induction of cell survival molecule NF-κB p65, and the phosphorylation of IκBα. It was also proven that fucoxanthin could repress the allergic rhinitis induced by OVA (albumin from chicken egg white). Benzoylpaeoniflorin is a cage-like monoterpenoid glycoside isolated from
Paeonia lactiflora extract [
58]. It shows anti-anaphylactic activity through the mechanism based on the inhibition of HDC (Histidine decarboxylase, crucial in histamine synthesis pathway) and MAPK signal pathways. Ginsenosides are active steroid compounds, first isolated in 1963 from ginseng. The studies have emphasized that ginsenoside Rh2 (G-Rh2) reduced calcium uptake and histamine release [
59]. The suggested mechanism included blocking IgE-induced degranulation by inhibiting AKT-NrF2 and p38MAPK-Nrf pathways. Studies of G-Rg3 have shown that it can decrease histamine release from MCs by enhancing cAMP levels and calcium influx, which is the opposite effect to Rh2. Protection against anaphylaxis is seen here in the regulation of mitogen-activated and receptor-interacting kinases in mast cells [
60]. Previous research showed that G-Rh2′s antiallergic activity can be exhibited by stabilization of cell membrane and inhibiting of NO and PGE2 production [
58,
61]. Some authors have also suggested that ginsenoside Rh1 possesses antiallergic activities. However, in this case, its effectiveness was sought in the inhibition of mast cell degranulation thanks to cell membrane-stabilizing and anti-inflammatory activities [
62]. Although studies have been conducted by many authors, the mechanism of ginsenosides is still insufficiently explored.
Another approach to managing anaphylaxis is focusing on Lyn kinase inhibition. Lyn kinase regulates phosphorylation of the protein scaffolds LAT (linker for activation of T cell) and NTAL (Non–T cell activation linker). Lyn is the first protein kinase phosphorylated and activated after allergen-IgE antibody complexes crosslink with FcεRI on the surface of MCs [
63,
64]. Alpha-linolenic acid is a substance that can modulate allergic reaction and MCs degranulation by inhibiting Lyn kinase activity, which was proven in vitro using LAD2 cells, and in vivo using OVA (albumin from chicken egg white)-stimulated mice model [
65]. A similar mechanism for preventing anaphylaxis is described in Ashikari’s article from 2022 [
66]. This time, salicylaldehyde was tested in vivo on the passive anaphylaxis mouse model. Studies have shown that the use of this substance reduced the symptoms of anaphylaxis in mice, such as increased temperature in the case of systemic anaphylaxis and swelling and vascular permeability in cutaneous anaphylaxis.
It is evident that the complex interactions between immune cells and structural cells in the inflammatory microenvironment determine how allergic reactions shift out. A similar situation is present in the microenvironment of cancer, where the immune system plays a crucial role [
46].
2. Mast Cell-Targeted Strategies in Cancer Therapy
For many years, mast cells were mainly known for their role in allergic reactions; however, the last decade has brought many new studies on these cells. The results clearly indicate that these cells also participate in the development of cancers, including hematopoietic cancers [
68,
69]. Mast cells play a multifaceted role in the tumor microenvironment by modulating various events of tumor biology, such as cell proliferation and survival, angiogenesis, invasiveness, and metastasis. Moreover, tumor-associated mast cells have the potential to shape the tumor microenvironment by establishing crosstalk with other tumor-infiltrating cells. MCs could stimulate the growth, neo-angiogenesis, and metastasis of tumors by multiple mechanisms [
70,
71,
72,
73].
Mast cells can exhibit anti-tumor activity either through: (a) direct tumor cell cytotoxicity and release of tumor necrosis factor or indirectly via mast cell-released heparin actions on fibroblasts, (b) acting as sentinel cells that secrete multiple chemokines that mobilize anti-tumor immune effector cells to tumor sites, (c) modulating immune effector cell responses and differentiation through the release of cytokines or through cell–cell interactions [
12,
72,
74].
On the other hand, MCs also exhibit pro-tumor effects. Activated mast cells can potentiate the deregulated tissue of the tumor microenvironment and favor tumor growth, and spread through: (a) the release of pro-angiogenic factors which enhance migration, proliferation, and blood vessel formation, (b) the release of proteases that release growth factors that have been sequestered in the (ECM) to enhance fibroblast proliferation and the angiogenic response and that degrade the ECM, thereby aiding tumor cell invasion of the, (c) contributing to the immune suppressive tumor environment through the release of cytokines such as TGF-β1 and IL-10 and indirectly through interactions with myeloid-derived suppressor cells (MDSC) and regulatory T (Treg) cells. Another known mechanism is tumor-derived TNF-α upregulating PD-L1 expression in the mast cells, representing a mechanism of immune suppression via the direct interaction between MCs and T lymphocytes in a PDL1-dependent manner [
12,
75].
Scientific studies confirm mast cell pro-tumoral function and the association of Tumor-Associated Mast Cells (TAMCs) with a poor clinical prognosis of various solid tumors. Mostly in the colon [
75,
76,
77,
78,
79], gastric [
80,
81,
82,
83], and pancreatic cancer [
84,
85,
86,
87], a growing number of clinical studies have associated high TAMC numbers with tumor progression and worse prognosis in patients. A similar association, though controversial, has been reported for breast [
75,
88,
89,
90,
91,
92], lung [
93,
94,
95], and prostate cancer [
96,
97]. Furthermore, recent studies showed that human melanoma-associated Mast Cells characterized by an upregulation of the complement component C 3 correlates with poor prognosis [
98].
The precise role of MCs in tumor development and progression will be crucial for the development of new targeted therapies in human cancers [
99]. The relationship between the MC density of tumors, the progression of angiogenesis, and tumor development may enhance the possible role of MCs in tumor biology. Therefore, the possibility of targeting MC activation [
100], inhibiting the release of mediators using c-Kit receptor tyrosine kinase inhibitors (TKI) (imatinib, mastinib, sunitinib [
99,
101]), or using tryptase inhibitors (gabexate mesylate and nafamostat mesylate, both inhibitors of trypsin-like serine proteases [
99,
102]) may be valuable therapeutic approaches to control the tumor development [
103].
Mastinib is an orally available inhibitor of the protein tyrosine kinase c-kit, which is expressed on the surface of cancer cells. Mastinib also inhibits PDGF and FGF receptors, and fyn and lyn kinases [
104,
105]. Furthermore, it has been used in veterinary medicine for years, and lately, human clinical trials were initiated to test its clinical efficacy as a single or add-on treatment for human cancers such as mastocytosis, gastrointestinal stromal tumors (NCT00998751), colon cancer (NCT03556956), prostate cancer (NCT03761225), and pancreatic cancer [
106].
Imatinib (“Gleevec” or “Glivec”), an orally available tyrosine kinase inhibitor, was called a “magical bullet” when it revolutionized the treatment of chronic myeloid leukemia (CML) in 2001. The first clinical trial of imatinib took place in 1998, and the drug received FDA approval in May 2001. The success of treating patients with CML with imatinib prompted scientists to investigate the therapeutic effect in other types of cancer, and it was found to produce a similar effect in other cancers where tyrosine kinase was overexpressed [
101]. Currently, imatinib is the standard of care in CML and GIST as it has dramatically changed the outlook of these diseases. It was the first cancer agent proven effective for metastatic GIST and represented a major development in the treatment of this rare but challenging disease. However, approximately 20% of patients do not respond to imatinib (early or primary resistance), and among those who do respond initially, 50% develop secondary imatinib resistance and disease progression within two years. Patients had no therapeutic option once they became resistant to imatinib before sunitinib was discovered [
107].
Sunitinib (“Sutent”) is a small-molecule, multi-targeted (RTK) inhibitor that was approved by the FDA for the treatment of renal cell carcinoma (RCC) and imatinib-resistant (GIST) on 26 January 2006. Sunitinib was the first cancer drug simultaneously approved for two different indications. It offers patients with imatinib-resistant GIST a new treatment option to stop further disease progression and, in some cases, even reverse it. This was shown in a large, Phase III clinical trial in which patients who failed imatinib therapy (due to primary resistance, secondary resistance, or intolerance) were treated in a randomized and blinded fashion with either sunitinib or placebo [
107].
Gabexate mesylate is a synthetic serine protease inhibitor that inhibits various kinds of plasma proteins, such as thrombin, plasmin, kallikrein, trypsin, C1 esterase in the complex system, and factor Xa in the coagulation cascade [
108,
109]. GM inhibits colon cancer growth, invasion, and metastasis by reducing matrix metalloproteinases. The antitumorigenic effect of GM is related in part to the antiangiogenic effect of GM [
110].
Nafamostat mesylate is a synthetic serine protease inhibitor; it is short-acting and is also used for the treatment of pancreatitis. Nafamostat also has some potential antiviral and anti-cancer properties [
111].
It seems that a combination chemotherapy of tryptase inhibitors or c-Kit receptor inhibitors and classical cytotoxic drugs could potentially exert a synergistic anti-tumor effect. Novel agents killing MCs might be evaluated in adjuvant clinical trials as a new anti-cancer approach.
In addition to the pro-tumoral functions, MCs may also modulate the response of cancer cells to therapy. The in vitro assays demonstrated that MC culture supernatants blocked gemcitabine (GEM)/nabpaclitaxel (NAB)-induced apoptosis in pancreatic cancer cell lines, through the activation of TGF-β1 signaling. Furthermore, these MC-derived supernatants reduced the anti-invasive activity of GEM/NAB. These data showed a functional interplay between MCs and pancreatic cancer cells, which induced resistance to GEM/NAB [
75,
86]. This observation was supported by the finding that unresponsiveness to GEM/NAB correlated with increased levels of tryptase and TGF-β1 in the blood of pancreatic ductal adenocarcinoma patients. Thus, MCs seem to play a crucial role in tumor resistance to GEM/NAB. Analysis of tumor tissue of inflammatory breast cancer (IBC), an aggressive form of breast cancer characterized by the clinical appearance of inflammation, showed that the MCs degranulating was significantly associated with poor response to neoadjuvant chemotherapy in all disease stages and molecular subtypes of IBC. Moreover, MCs were located within range for direct or paracrine interactions with CD8
+ T cells, as well as CD163
+ macrophages and tumor cells. The authors suggested that the interaction of MCs with these immune cells might be exerting an inhibitory effect in IBC, through suppressing CD8
+ T cells, enhancing immunosuppressive CD163
+ macrophages, and directly promoting tumor cell growth [
75,
92]. This study indicated that MCs could represent a possible therapeutic target to enhance the response to chemotherapy.
This entry is adapted from the peer-reviewed paper 10.3390/cells12081187