]. The Li
cation is therapeutically relevant, given that it has been used to treat bipolar disorders [
]. For bipolar mood disorders, achievable Li levels in plasma are around 0.5–1.0 mM (mEq/L) [
], whereas concentrations higher than 2 mM cause cytotoxicity in patients, resulting in nausea, fine tremors, diarrhea, confusion, slurred speech, and ataxia [
]. Lithium forms a complex with ATP and Mg
, resulting in the bioactive form of lithium that modulates purine receptor activity in neuronal cells [
]. Lithium had been used over decades for bipolar disorder treatment, preventing relapses, and for the prophylaxis of manic episodes [
]. Some morphological abnormalities have been observed in patients with bipolar disorder. The neuroprotective activity of Li is related to the inhibition of its molecular targets, such as the enzymes inositol-monophosphate (IMPase), glycogen-synthase-kinase 3β (GSK3), and protein kinase C (PKC) [
]. Interestingly, these enzymes are deregulated in cancer, suggesting a potential role for Li, beyond bipolar disorders, in cancer treatment. Recent in vitro and in vivo studies reported the effects of lithium (lithium acetoacetate (LiAcAc), lithium chloride (LiCl), lithium citrate (Li
) on cell proliferation, tumor growth, and cancer metabolism. According to this evidence, LiAcAc inhibited cell proliferation in colon, breast, pancreatic, and neuroblastoma cancers [
].
The Li
+ ion shares transport and permeation pathways with other ions, such as sodium [
38]. Therefore, extracellular and intracellular concentrations of the Li
+ ion depend on its ingestion [
39]. However, LiAcAc requires specific transporters. In cells, the presence of monocarboxylate transporters (MCTs) is involved in the passive transport of lactate, pyruvate, D-β-hydroxybutyrate, and acetoacetate across the cell membrane [
40]. MCT2 has the highest affinity constant (Km) for KBs such as D-β-hydroxybutyrate and acetoacetate [
41], but these substrates also are transported by MCT1 [
42] and MCT4, which have a lower affinity for them [
43]. LiAcAc is imported into cells through MCT1 and MCT2, with Km values of 5.5 mM and 0.8 mM, respectively [
41,
43,
44]. These values indicate that MCT2 has a higher affinity for LiAcAc compared to MCT1, therefore the cellular expression of MCT2 could determine the cellular response to Li
+ ion exposure. Differential expression of MCTs in healthy and cancer tissues has been reported. In this way, MCT1 is expressed by healthy or cancer tissues [
45]. Furthermore, MCT1 has a specific expression in cardiomyocytes, hepatocytes, and cells of the gastrointestinal tract (stomach, duodenum, jejunum, ileum, cecum, colon, and rectum) [
44,
46,
47,
48,
49,
50,
51,
52,
53,
54,
55]. In addition, this transporter is found in erythrocytes, lymphocytes, and monocytes [
46,
56], in adipocytes [
57,
58] and in mammary glands [
59]. Moreover, MCT1 is found in endothelial brain cells, astrocytes, oligodendrocytes, microglial cells, and the hypothalamus [
60,
61,
62,
63,
64]. Furthermore, this transporter was found in the peripheral nervous system [
65,
66]. While MCT1 is a highly distributed transporter, MCT2 has a restricted distribution, being found in the liver and kidney, testis, and central nervous system [
44,
67,
68,
69,
70]. The ratio of intracellular/extracellular lithium concentration is about five, suggesting a passive distribution of Li
+ ion. The transport of lithium is in both directions across the cellular membrane through a Na
+-dependent counter-transport system. Furthermore, Li
+ ion enters the voltage-dependent Na
+ channel, but the Na
+-K
+ pump mediates the lithium uptake but not its release in cells [
71]. Therefore, neuronal cells maintained a ratio of internal-external Li
+ ion below 1 mM in patients exposed to 1–2 mM of lithium [
71].
Lithium Transporters in Cancer
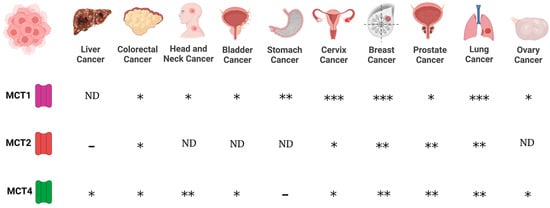
Regarding MCTs, evidence indicates that MCT1 and MCT4 are expressed in cancer cells, whereas scant information is available regarding MCT2 and MCT3 expression in cancer. Expression of MCT1 was reported for lung (non-small cell lung carcinomas, NSCLC), stomach, colon, bladder, prostate, breast, ovarian cervical, head and neck cancers, and gliomas [
72,
73,
74,
75,
76,
77,
78]. On the other hand, MCT4 is expressed in colon, bladder, and breast cancers, and gliomas [
74,
76,
77,
78]. In addition, while MCT2 is expressed in NSCLS, colon, breast, and ovarian cancers [
74,
78], MCT3 is found in NSCLC [
78] (
Figure 1).
Figure 1. Expression of monocarboxylate transporters (MCTs) in various cancers. MCTs are involved in the passive transport of lithium acetoacetate across cell membranes, and their abundance is associated with cancer. (-), no expression; (*), medium expression; (**), high expression; (***), very high expression level; ND, not data.
Furthermore, chaperone proteins of MCTs are important mediators in cancer. CD147 and CD44 function as chaperones of MCT1. In this way, the expression of MCT1 and CD147 correlated in bladder and ovarian carcinomas [
78], whereas the expression of CD44 is associated with MCT1 expression in lung cancer [
74].
4. Lithium as a Specific Inhibitor of GSK3β
The chlorinated form of lithium inhibits Glycogen Synthase Kinase 3β (GSK3β), which is a serine/threonine protein kinase. GSK3β phosphorylates β-catenin in the Wnt metabolic pathway, leading to growth arrest [
94,
95]. Moreover, GSK3β participates in cellular proliferation and survival by activating nuclear factor ϰB-dependent gene transcription, suggesting an active role of GSK3β in tumorigenesis in pancreatic, colorectal, and prostate cancers, and gliomas [
96,
97,
98,
99,
100]. Since GSK3β is involved in repressing Wnt/β-catenin signaling, but also in maintaining cell survival and proliferation via the NF-ϰB pathway, the inhibition of this dual kinase activity by lithium might have a positive effect on cancer hallmarks.
The expression of GSK3β is higher in colon cancer cells and in colorectal cancer patients in comparison to normal counterparts. Furthermore, the inhibition of GSK3β activity resulted in apoptosis induction and reduced proliferation in cell culture [
99]. Interestingly, lithium is capable of inhibiting GSK3β; consequently, lithium is a potential therapeutic agent. Inhibition of GSK3β by LiCl takes place through two mechanisms. One of them is a competitive mechanism between lithium and magnesium for binding to GSK3β [
101,
102]. The other mechanism involves the role of LiCl in the phosphorylation of the serine-9, located in the N-terminal, which is the main regulator region of GSK3β [
103]. However, GSK3β has contradictory functions in cancer cells. On one hand, GSK3β phosphorylates p53, leading to its inhibition [
104]. In this way, GSK3β binds to p53 in camptothecin-treated neuroblastoma cells, leading to increased mitochondrial apoptosis signaling and the expression of p21 and Bax [
105,
106].
Furthermore, when lithium blocks GSK3β activity, ovarian cancer cell proliferation was reduced and tumor suppressor activity was observed in nude mice inoculated with human ovarian cancer cells [
108]. Moreover, proliferation was reduced in Li-treated human medullary thyroid TT cancer cells, due to the inactivation of GSK3β activity [
109].
5. Effect of Lithium on Certain Cancer Hallmarks
5.1. Effect of Lithium on Apoptosis
The role of lithium as a proapoptotic or antiapoptotic compound is contradictory. In HepG2 cells, lithium confers resistance to the apoptosis induced by etoposide and camptothecin, by preventing the activation of caspase-8 and caspase-3 and by inhibiting the Bax translocation to mitochondria [
107]. Therefore, lithium confers resistance to chemotherapy-induced apoptosis through GSK3β inhibition, disruption of GSK3β/p53 cooperation, and repression of CD95 expression [
107].
On the other hand, in human SH-SY5Y neuroblastoma cells, lithium inhibits GSK3β, contributing to antiapoptotic signaling mechanisms [
113]. Furthermore, LiCl induces the decrease of proapoptotic proteins p53 and Bax, while increasing Bcl-2 (a major antiapoptotic protein that controls mitochondrial membrane permeability) levels [
114]. In addition, the effects of mitochondrial toxins might be attenuated by the inhibition of GSK3β, which can be achieved by the activation of the phosphatidylinositol 3-kinase (PI3K)/Akt pathway, which is inhibited by GSK3β through Akt-mediated phosphorylation of Ser-9 of GSK3β [
115] or by GSK3β direct inhibitors such as lithium, suggesting that the inhibition of GSK3β provides protection against the activation of the apoptosis-associated cysteine protease caspase-3 [
116].
In glioblastoma cells, low and higher LiCl doses had no effect on the expression of Bcl-2, procaspase-3, and PARP [
118]. In contrast, in neuronal cells, Li blocks caspase-3 activation [
119]. Therefore, cells of different origins (neuronal or glial) might have different results when treated with lithium. In cancer, several pathways might be dysregulated, and combinatory therapy with LiCl and pharmacological activators of the Notch1 pathway suppressed the hormonal secretion and reduces cell growth through apoptosis in medullary thyroid cancer (MTC) [
120]. Lithium chloride enhances cell death induced by tumor necrosis factor-related apoptosis-inducing ligand (TRAIL) in human prostate cancer cells [
121].
5.2. Effect of Lithium on Autophagy
Lithium induces autophagy and enhances the clearance of aggregated proteins, such as mutant huntingtin and α-synucleins neuronal precursor cell lines and non-neuronal cells. Interestingly, this effect is not mediated by GSK3β, but it was mediated by inhibition of IMPase, leading to free inositol depletion and reducing myo-inositol-1,4,5-triphosphate (IP
3) levels, resulting in a novel form of autophagy induction that is independent of the mammalian target rapamycin (mTOR) [
127,
128]. In addition, the inhibition of IMPase and GSK3β showed opposite effects on autophagy. On one hand, Li induced the inhibition of IMPase at lower doses (Ki = 0.8 mM), enhancing autophagy [
128].
Additionally, lithium carbonate promotes autophagic vacuole formation in hepatocellular carcinoma (HCC) cells in vivo, suggesting that lithium-mediated autophagy might be a novel approach for treating hepatocellular carcinoma [
133]. Interestingly, in HCC, combinatory treatment of lithium carbonate and rapamycin increases autolysosome formation and the expression of autophagy markers such as LC3β and LAMP1 [
134]. Further, 5 mM lithium carbonate produced an antitumor effect, by arresting the cell cycle in G2/M-phase and increasing the number of apoptotic HCC cells, as well as inducing autophagy [
33]. In corneal endothelial cells, treatment with lithium carbonate upregulated the expression of
P62,
Tmem74,
Tm9sf1, and
Tmem166, indicating that lithium increased autophagy that may have contributed to increased endothelial cell survival in a mouse model of Fuchs endothelial corneal dystrophy [
135].
5.3. Effect of Lithium on Tumor Growth, Tumor Proliferation, Tumor Invasion and Metastasis, and Cell Cycle Arrest
Tumor cell growth is controlled by various signaling pathways, such as PI3K/Akt and the raf-1/mitogen-regulated extracellular kinase (MEK)/extracellular regulated kinase (ERK) pathway (raf-1/MEK/ERK). LiCl decreases hormonal secretion and suppresses the growth of MTC in vitro and in vivo due to GSK3β inhibition [
109]. The raf-1 activation induces GSK3β phosphorylation, therefore the inactivation of GSK3β by LiCl might be a mechanism of rat-1-induced growth arrest [
109].
Interestingly, low concentrations of LiCl (at 10, 20, 30, and 40 mM) promote the proliferation of mk3 and mk4 cells (metanephric mesenchyme cells), which play an essential role in nephron generation, by increasing the expression of the transcription factor
Six2 (
sine oculis homeobox homolog 2 gene). In contrast, higher concentrations of LiCl (50 mM) inhibit proliferation and downregulate the expression of
Six2 [
141].
Lithium causes nuclear localization of the claudin-1 and β-catenin that form the apical junctional complex, which is associated with colorectal cancer progression. In addition, lithium reduces colony formation, inhibiting cell migration and cell proliferation through G2/M cell cycle arrest. In colorectal cancer, the aberrant activation of epidermal growth factor (EGF) disrupts the association between E-cadherin and β-catenin, increasing cell proliferation and migration [
1]. EGFR activates downstream kinases that modulate the GSK3β phosphorylation and consequently activate the Wnt/β-catenin pathway [
145,
146]. Since LiCl inhibited the tumorigenic effects of EGF, this cation might have tumor suppressor activity in colorectal cancer [
147]. LiCl inhibits multiple-myeloma proliferation in a dose-dependent manner and triggers cell cycle arrest and apoptosis through inhibition of GSK3β, which is a crucial mediator of the Wnt/β-catenin pathway [
24]. GSK3β regulates cadherin-11 at transcriptional and translational levels. Lithium inhibition of GSK3β suppresses cadherin-11, which is involved in cell–cell adhesion, cancer cell invasion, and metastasis of cancer [
148].
In colon cancer, lithium reduces the expression of transforming growth factor-β-induced protein (TGFBIp) by inhibiting Smad3 phosphorylation through GSK3β inhibition [
150]. In addition, Li inhibits cell migration of lymphatic endothelial cells and prevents metastasis by inhibiting TGFBIp-induced tumor lymphangiogenesis, resulting in the inhibition of colon cancer metastasis [
150]. The effects of lithium on migration are greater than its effects on viability and occur at lower doses and earlier time points. Moreover, the migration effects of Li are reversible, suggesting that migration blockade is not due to cytotoxicity [
151].
6. Anti-Inflammatory Activity of Lithium
In some types of cancer, before the cell becomes malignant, inflammatory conditions occur. Therefore, inflammation is associated with the proliferation and survival of cancer cells. In addition, this promotes angiogenesis and metastasis [
159]. Since inflammation is associated with the development and progression of cancer, inflammatory biomarkers such as pro-inflammatory cytokines, tumor necrosis factor (TNF), interleukins 1 and 6, and chemokines can be used to monitor cancer progression [
160]. In this way, the anti-inflammatory effect of lithium might be an alternative for prevention and monitoring of cancer. Furthermore, in animal models, LiCl treatment induced therapeutic effects on inflammation-mediated skeletal muscle wasting such as sepsis-induced muscle atrophy and cancer cachexia [
161].
The anti-inflammatory activity of lithium results from its strong inhibitory effect on GSK3β, leading to a reduction in TNF-α production via attenuated activation of NF-κB and JNK signaling cascades [
124] and induction of cytokine IL-10 [
162]. The anti-inflammatory activity of lithium is due to the reduction of cyclooxygenase-2 expression, induction of the nitric oxide synthase expression, inhibition of interleukin 1β and TNFα, and increased production of IL-2 and IL-10 [
163,
164].
7. Synergism of Lithium with Standard Cancer Therapies
High concentrations of lithium (4–5 mM) inhibit inositol 1-monophosphatase, resulting in a reduction of phosphatidylinositol turnover. However, these high concentrations of Li
+ ion have toxic effects in patients [
14]. Reported serum Li
+ ion levels for therapy were 0.8 to 1.2 mEq/L (0.8–1.2 mM) and the cytotoxicity effect began at 1.5 to 2 mEq/L (1.5–2 mM) [
167]. Interestingly, 1 mM of LiCl had no effect on cellular metabolism or cell cycle but its combination with cisplatin or paclitaxel reduced metabolic activity in a serous ovarian cancer cell line (OVCA 433) or primary cultures. However, LiCl by itself or in combination with cisplatin or paclitaxel had no significant effect on cellular proliferation, suggesting that the effects of LiCl are mainly related to cellular metabolism rather than proliferation [
167].
Conventional cancer therapies such as chemotherapy and radiotherapy inhibit cell division, causing neurocognitive deterioration. Preliminary studies have shown that lithium has a neuroprotective role in cancer patients undergoing radiotherapy or chemotherapy treatment, improving their quality of life [
169]. Combination of LiCl with a low dose or IC
50 dose of etoposide enhances apoptosis and reduces the percentage of cells in the G2/M phase in prostate cancer LNCap cells [
19]. In pancreatic ductal adenocarcinoma cells, lithium increases the anti-tumoral efficacy of gemcitabine [
125]. Additionally, histone deacetylase inhibitors with lithium resulted in an effective combinatory therapy, due to inducing the Notch1 pathway and inhibiting GSK3β activity, respectively. [
120].
Chemotherapy-induced peripheral neuropathy (CIPN) is an adverse effect of cancer treatment using taxanes, such as paclitaxel. Adding lithium to taxane treatment prevents calcium overloading in mitochondria, resulting in the prevention of calpain activation and preventing the loss of cell functions related to the progression of peripheral neuropathy [
177], suggesting that lithium could be used to prevent the adverse effects of cancer treatment with only taxane administration [
178].
8. Contradictory Effects of Lithium
Prolonged treatment with lithium causes renal, endocrine, and dermatological side effects. Four weeks of therapy with lithium caused renal tubular concentration defects, polyuria, polydipsia, and nephrogenic diabetes insipidus [
208]. Administration of Li
2CO
3 induced a cutaneous adverse reaction [
209] and parathyroid dysfunction [
210]. Furthermore, lithium treatment caused a higher prevalence of hypercalcemia in a cohort of bipolar patients [
211]. In addition, there is a possibility of teratogenicity activity with lithium treatment [
212].
The inhibition of GSK3β using lithium has contradictory effects. On one hand, chemoresistance to apoptosis is observed, but on the other hand, it induces tumor suppressor activity and reduces proliferation. Those effects depend on the type of cancer, the origin of cells, and doses. The anti-cancer effect of lithium has been reported for colorectal cancer, neuroblastoma cells, ovarian cancer cells, medullary thyroid TT cancer cells, esophageal cancer, medulloblastoma, and glioblastoma multiforme, hematological tumors (multiple myeloma), breast cancer cells, MTC, prostate cancer cells, pancreatic ductal adenocarcinoma cells, corneal endothelial cells, and colon cancer cells. However, opposite effects of lithium have been reported for neuroblastoma, hepatoblastoma cells, and malignant glioma cell lines.
Lithium induces apoptosis in promyelocytic leukemia cells, but a small fraction of these cells remain viable and survive the cytotoxic effect of this cation, resulting in altered gene expression patterns [
213], suggesting cell tolerance to lithium in culture [
214]. Resistance to lithium is a consequence of its activity. Lithium appears to induce apoptosis by increasing the
bax/
bcl-2 ratio (
bax: pro-apoptotic gene and
bcl-2: anti-apoptotic gene). Lithium resistance then results when this ratio is reversed, leading to lithium resistance accompanied by DNA fragmentation, and decreasing cell viability [
158].