The aryl hydrocarbon receptor (AhR) is a ligand-activated transcription factor involved in regulating a wide range of biological responses. A diverse array of xenobiotics and endogenous small molecules bind to the receptor and drive unique phenotypic responses. Due in part to its role in mediating toxic responses to environmental pollutants, AhR activation has not been traditionally viewed as a viable therapeutic approach. Nonetheless, the expression and activation of AhR can inhibit the proliferation, migration, and survival of cancer cells, and many clinically approved drugs transcriptionally activate AhR. Identification of novel select modulators of AhR-regulated transcription that promote tumor suppression is an active area of investigation. The development of AhR-targeted anticancer agents requires a thorough understanding of the molecular mechanisms driving tumor suppression.
1. Role of AhR in Xenobiotic Metabolism
AhR belongs to the superfamily of transcription factors containing both basic helix–loop–helix (bHLH) and PER-ARNT-SIM (PAS) domains (14). The PAS domain is conserved across all kingdoms of life, and functions in sensing and activating biological responses to environmental changes. Related transcription factors containing the PAS domain, such as hypoxia-inducible factor 1α (HIF1α), and circadian clock proteins such as PER1 and BMAL, respond to environmental cues such as oxygen tension and light [
14]. In this respect, the aryl hydrocarbon receptor functions as both an internal and external sensor of chemical signals.
The role of AhR in the cellular detoxification or metabolism of small molecules is well established [
15]. Exposure to xenobiotics, or activation by endogenous metabolites, induces the receptor’s activation and a subsequent transcriptional response that then attempts to bring the cell back to a state of homeostasis. A major component of this homeostatic response is induction of the cytochrome P450 enzymes, including CYP1A1, CYP1A2, and CYP1B1, which function, through substrate oxidation, to convert the ligands to more water-soluble forms that can be conjugated by Phase II enzymes (e.g., glutathione-S-transferase) and excreted via Phase III enzymatic transport [
16]. While it was evolutionarily acquired as a transcriptional circuit to ameliorate xenobiotic insults, exposure to certain xenobiotics can induce AhR-mediated toxicity. It is in this context that AhR has been historically and extensively studied, as AhR was originally cloned and identified as the receptor responsible for mediating the biological effects of dioxin (2,3,7,8-tetrachloro-dibenzo-
p-dioxin (TCDD)) [
17], a legacy anthropogenic pollutant. TCDD binds with high-affinity to AhR, is resistant to CYP1-mediated metabolic degradation, and is highly lipophilic. Together, these properties make TCDD highly bio-accumulative, and its toxic effects are from chronic AhR activation [
18].
Within an organism, activation and antagonism of AhR have diverse consequences that depend critically on the cellular abundance and characteristics of the initiating ligand, as well as specific tissue and organ dynamics. While AhR is a master regulator of xenobiotic metabolism, it also has important roles in processes such as development, differentiation, and immune function.
2. The AhR Signaling Pathway
The transcriptionally inactive AhR complex is located in the cytosol, sequestered by an HSP90 dimer [
19,
20], co-chaperone p23 [
21,
22], AhR-interacting protein (AIP), and the protein kinase Src [
23]. The complex functions to both sequester AhR in the cytosol and hold the AhR in conformation to interact with ligands [
24]. Upon an agonistic ligand binding to the receptor, AhR-associated AIP dissociates from the receptor, exposing its nuclear localization sequence, and importin-β then binds to AhR and transports the receptor into the nucleus.
Once in the nucleus, AhR can then interact with the heterodimer partner, aryl hydrocarbon receptor nuclear translocator (Arnt), also known as HIF-1β, where the AhR–Arnt complex then binds to the regulatory regions of DNA containing the consensus motif (5′-TNGCGTG-3′) [
25,
26] or the xenobiotic response element (XRE) (also known as dioxin response element (DRE), or the AhR response element (AhRE) and modulates the transcription of the target gene. While XRE-driven transcription through AhR–Arnt signaling represents the most well studied mode of regulation, AhR can modulate gene expression through non-XRE elements [
27,
28] and can also interact with other transcription factors including c- MAF [
29], KLF6 [
30], RelA [
31], and other NF-κB complex members [
32].
Structurally, AhR contains the following features, proceeding from its amino terminus: a bHLH domain (amino acids (a.a.) 33–87); PAS-A (a.a. 111–273) and PAS-B domains (a.a. 275–386); and a transactivation domain (TAD) that contains an acidic region, a glutamine-rich (Q-rich) region, and a proline/serine-rich region (a.a. 490–805) [
33,
34]. AhR binds to the regulatory regions of DNA via its bHLH domain [
33], which is masked by one molecule of HSP90 in the unliganded cytoplasmic complex [
19], partially overlapping with the PAS-A region. The second molecule of HSP90 interacts dominantly with the PAS-B region. The bHLH domain dimerizes with the bHLH of Arnt to form a bundle of four helixes that interact with the XRE [
25]. Similarly, the PAS-A domains of AhR and Arnt function as a dimerization interface, conferring specificity and enhancing the complex’s stability and DNA binding [
35,
36]. The PAS-B domain functions as the ligand-binding domain [
36] and is required for nuclear translocation of the receptor, but is dispensable for heterodimerization and ligand-binding [
37,
38]. Notably, high-affinity ligand-binding and the subsequent induction of metabolizing enzymes represents a vertebrate adaptation and has been suggested as an acquired function in response to aromatic marine natural products [
1]. Early invertebrate AhR paralogs in
Caenhorabiditis elegans [
39,
40] and
Drosophila melanogaster [
41] function primarily in neuronal and sensory development, and this role is retained in vertebrates in addition to its ligand-mediated activities [
42].
Species-specific [
43], ligand-specific [
44,
45], dose-dependent [
46], tissue-specific, and microenvironment-specific factors [
47,
48] together influence the apical biological outcome downstream of the activation of the AhR ligand, so making initial predictions of a ligand’s effects in a given model system is extremely difficult. For example, species-specific divergences in the receptor’s structure and ligand-binding affinity result in significant differences in the transcriptional [
43] and phenotypic response to ligands such as TCDD between humans [
43], mice [
49], and guinea pigs [
50]. The mouse AhR differs from its human ortholog in its ligand-binding affinity due to a single amino acid substitution in the ligand-binding pocket [
51]. Consequently, TCDD binds with significantly greater affinity (approximately 10×) to the mouse AhR, while distinct ligands such as indoxyl-3-sulfate (of endogenous origin) bind with greater affinity to the human receptor [
51,
52]. To understand some of these interspecies differences, our laboratory previously generated an AhR homology model for the AhR PAS domain [
53,
54].
3. Role of Co-Regulators in AhR Signaling
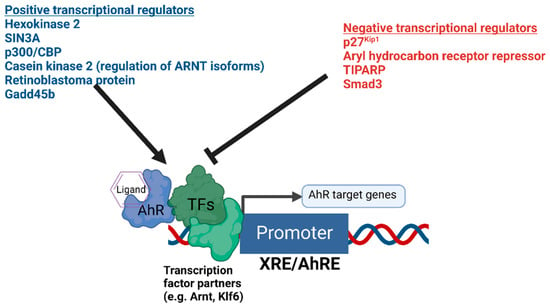
The presence or absence of co-regulatory proteins and their activity levels influence both the basal transcription and ligand inducibility of AhR [
55,
56] (
Figure 1). Recently, hexokinase 2 was found to be a transcriptional target of AhR and a positive regulator of AhR-mediated transcription [
57]. Important regulators of AhR signaling also include proteins such as AhRR [
58], TiPARP [
59,
60,
61,
62], Gadd45b [
63], p300, CREB binding protein (p300/CBP) [
64], Smad3 [
65], and SIN3A [
66,
67]. AhRR and TiPARP function to provide negative feedback on AhR’s transcriptional activity, while SIN3A, typically regarded as a transcriptional repressor, was found to function as an essential co-activator for the transcription of CYP1A1 [
67]. An additional mechanism fine-tuning the function of AhR was recently identified [
68]. Bourner et al. found that differential expression levels of ARNT isoforms altered the transcriptional activity of AhR, and ARNT isoform-dependent regulation of AhR was shown to be dependent on phosphorylation by the AhR target gene casein kinase 2 [
68].
Figure 1. Summary of the positive and negative transcriptional co-regulators of AhR signaling. The diagram depicts AhR bound to DNA in complex with potential heterodimer partner proteins and regulators. Different positive and negative regulators of AhR-mediated transcription are summarized above.
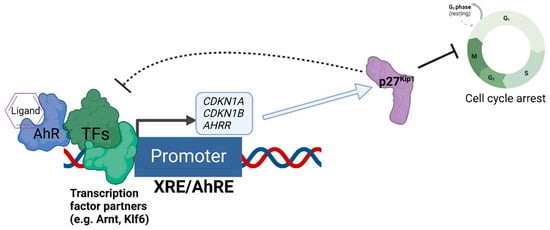
We recently discovered that the expression of the cell cycle inhibitor and direct AhR target gene, p27
Kip1, represses AhR-mediated transcription [
69] (
Figure 2). Loss of p27
Kip1 in lung cancer cells resulted in significantly enhanced basal and ligand-induced levels of AhR target genes such as CYP1A1 and AHRR, and this negative regulation was found for the transcription of p27
Kip1 itself [
69]. Interestingly, p27
Kip1-dependent transcriptional repression was previously found to operate via the formation of a protein complex containing mouse retinoblastoma protein (Rb) homolog p130, mSIN3A, E2F4, and multiple histone deacetylases (HDACs 1, 4, and 5) [
70,
71], Further, AhR’s interaction with Rb [
72] and transcriptional regulation of AhR downstream from Rb have been reported [
73], indicating bidirectional regulation between AhR and components of the cell cycle’s machinery.
Figure 2. Negative regulation of AhR-mediated transcription by the expression of p27Kip1. The diagram depicts the regulatory relationship between p27Kip1 and AhR, whereby AhR acts as a direct transcriptional regulator of the transcription of p27Kip1, and increased expression of p27Kip1 both inhibits cell cycle progression and represses the AhR-mediated transcription of genes such as CDKN1A (encoding p21Cip1), AHRR, and CDKN1B (encoding p27Kip1 itself) as an auto-feedback loop.
The p300/CBP enzymes function as transcriptional co-activators via their acetyltransferase activity, and the catalytic activity of p300/CBP was found to be essential for AhR’s transcriptional activity [
64]. Notably, ARNT was identified as the key target of p300/CBP’s acetylation for activating transcription [
64]. Interestingly, previous reports indicated that the acetylation of the p27
Kip1 by p300/CBP-associated factor (PCAF) attenuated its transcriptional repressor activity and promoted its degradation, suggesting an additional layer of regulatory control [
74,
75].
This entry is adapted from the peer-reviewed paper 10.3390/biology12040526