1. Essentials of mRNA Vaccines Design and Functioning
Although various mechanisms of infectivity have been described [
45,
46,
47], entry into cells by SARS-CoV-2 relies mainly on the interaction of the envelope Spike protein with cellular ACE2. Thus, blocking this interaction with an antibody seemed a good strategy. This prompted the vaccine production industry to design a genetically engineered vaccine capable of inducing in the host the production of neutralizing antibodies against the Spike protein, particularly toward the interacting region called the RBD. Indeed, the immunizing antigen is produced within the hosting cell once the nanoparticle load of mRNA is injected. Thus, for the vaccine to trigger the immune response, i.e., to elicit the biological (immunological) effect, the mRNA must be translated into the protein that, in turn, must interact with the immune system.
The conception and rapid production of these new vaccines against SARS-CoV-2 followed within a few months after the Chinese authorities disclosed the sequence of the virus isolated in Wuhan. Western pharmaceutical companies rushed to use this sequence, and in particular the RNA “message” encoding the Spike protein, using a technology that was already available [
12,
48] yet never exploited on a large scale for human use. Seneff et al. [
49] carefully and extensively analysed many of the critical points related to the engineered mRNA vaccines.
The vaccine mRNA was engineered to increase its stability, to escape cellular degradation, and to ensure the production of the Spike protein with the RBD accessible for inducing neutralizing antibodies [
13,
14]. It is noteworthy that the mRNA vaccine sequence maintains the furin cleavage site (a stretch of the four basic amino acids Arg- Arg- Ala- Arg at the S1–S2 junction) as in the viral sequence, and this has implications for the generation of the soluble S1 peptide [
14,
22].
The original sequence of the protein was slightly modified (i.e., K986 and V987 in the S2 subunit were substituted by two prolines) to direct the synthesis of the protein in a stabilized “pre-fusion” (open) conformation, like that interacting with the ACE2 cell receptors and to which neutralizing antibodies are supposed to react [
13,
14]. Other modifications are briefly described below. To allow entrance into the cells, the mRNA is encapsulated in lipid nanoparticles (LNPs) containing cholesterol and phospholipids associated with modified polyethylene glycol to avoid its degradation [
50]. Viral RNA is recognized by the human cells as foreign, and this triggers defence reactions that impair its translation into proteins, while directing its degradation [
51,
52]. Replacing uridines with pseudouridines or (even better) with methyl-pseudouridine, overcomes the recognition as a foreign mRNA by the Toll-Like Receptors (TLR) and the subsequent activation of IFN type I [
53]. To stabilize the mRNA and thus improve its translation, anti-COVID-19 mRNA vaccines have this characteristic [
54]. To further stabilize the mRNA and increase the S protein production, a long poly(A) tail [
55] and the 3′ UTR from human globin [
56] were added to the mRNA molecule. A leader sequence, for translation in endoplasmic reticulum associated ribosomes, was added to ensure the insertion of the Spike protein into the plasma membrane. Notably, mRNAs vaccines are enriched in GC content: 53% in BNT162b2 and 61% in mRNA-1273 compared to 36% in native SARS-CoV-2 mRNA [
57], and this also contributes to increasing the protein production [
58]. Taken together, the vaccine mRNAs driving the Spike protein synthesis have been engineered in a manner that challenges the cellular stress response for the recognition of exogenous nucleic acids and proteins, and this is likely to impact the distribution of the mRNAs coding for the Spike protein and of the protein itself, which may then explain the biological and pathophysiological effects in organs distant from the site of injection. Indeed, the true biodistribution and the half-life of the vaccine mRNA in humans are currently unknown. Normally, mRNA is very fragile and is quickly degraded (within a few days). It was initially thought that vaccine mRNA would remain localized in the site of injection and be degraded within a few days, as is normal mRNA. However, real-world observations contradict this prediction. The S-protein has been detected in the plasma of mRNA-1273 COVID-19 vaccinees at 15 days following injection [
59]. Both mRNA and S protein have been found in axillary lymph nodes after 60 days [
60]. Very recently, Spike-mRNA has been detected in the blood of vaccinated individuals 15 and up to 28 days after COVID-19 vaccination [
61,
62]. Thus, it is likely that mRNA-LNPs remain in circulation for extended periods of time, retaining their ability to induce S protein expression in encountered cells. Updated bivalent mRNA vaccines that include the coding sequence for the Omicron BA.4/BA.5 variant were made available in September 2022, and studies on their efficacy and safety are still ongoing. Based on two pre-print studies, not yet peer-reviewed, the bivalent mRNA vaccine shows modest protection [
63] and a higher rate of adverse events compared to the monovalent mRNA vaccine [
64].
2. The Immune Response to the SARS-CoV-2 and to the mRNA Vaccines
The exact mechanism of stimulation of the immune system by the Spike protein-encoding vaccines is still hypothetical, and several versions exist. The first problem concerns the interaction of the injected product with the host.
2.1. The Importance of the Route of Entry
Normally, pathogens enter the body via different routes, namely the oral and gastrointestinal mucosa, the nasal mucosa, the urogenital mucosa, and the skin. Each of these routes is characterized by a peculiar local microenvironment (stromal cells, tissue-specific factors, and commensal microbiota) which heavily influences the type and extent of the innate and specific immune response. When an infectious agent, a toxin, or a foreign antigenic molecule enters the body tissues or the blood, the immune system mounts a robust proinflammatory response, involving first the innate (non-antigen specific) immune system and, if required (depending on the type of antigen, route of entry and its persistence), the adaptive antigen-specific immune system.
With evolution, the immune system has become more and more compartmentalized (cutaneous immune system, mucosal immune system, and systemic immune system) to improve its response and to reduce the risk of a dysregulated and disproportionate reaction. At the same time, the cells of the immune system can travel between the compartments and be influenced by the different local environments. The compartmentalized immune tissues communicate with each other to alert the system to the presence of the foreign potentially harmful “enemy” via the release of exosomes containing informative molecules (cytokines, microRNAs, PAMPs (Pathogen Associated Molecular Patterns), DAMPs (Damage Associated Molecular Patterns) from APCs and phagocytic cells [
65]. Notably, circulating exosomes with inserted on the membrane the Spike protein have been detected in vaccinated individuals, and it is assumed that such exosomes are internalized by the APC, thus adding another route of immune sensitization [
66].
The anatomical compartment determines the characteristics (differentiation status, phenotype, function, duration, turnover rate, homing capacity and regulatory mechanisms) of the immune cells. The threshold for activating the immune system is different in each organ and correlates inversely with its relative sterility [
67,
68]. To obtain effective and long-lasting protection at the site of entry, the pathogen must have direct contact with and be processed by the local tissue and compartmentalized immune system [
67].
As with other respiratory viruses, in the case of SARS-CoV-2 infection, the early phase of humoral response is mediated by IgA antibodies that show greater neutralizing activity than IgG [
69]. Upon viral infection, plasmablasts with homing receptors for mucosal sites and with intracellular IgA increase in the blood [
69]. The fact that the level of secretory IgA specific for the Spike RBD in saliva was higher than that in the blood of the same subject 49 days after the onset of symptoms is indicative of the persistence of IgA in the oral mucosa [
69].
The dimeric form of IgA, found in all secretions of both respiratory and intestinal mucosa, against SARS-CoV-2 is more potent than the monomeric IgA [
70,
71]. Salivary IgA specifically for the Spike protein are significantly lower in anti-COVID-19 mRNA vaccinees than in COVID-19 convalescent controls [
72]. In fact, the current mRNA vaccines, though able to prevent/attenuate the most serious consequences of the disease, do not trigger the mucosal IgA response [
73], even after the booster [
74], and do not prevent the colonization of the virus in the mucous membranes [
16]. The pattern of the cytokine response is also of paramount importance. The immune response to the virus and to the mRNA vaccines differ in that the former is characterized by strong induction of interferon and circulating effector B and T lymphocytes, whereas the latter is essentially restricted to circulating memory cells [
75].
2.2. Immunization Pathways of the SARS-CoV-2 and mRNA Vaccines
mRNA COVID-19 vaccines are meant to induce B lymphocytes capable of producing antibodies against the (viral) S protein for preventing SARS-CoV-2 entry into the cells as well as T lymphocytes capable of killing the virus-infected cells (in the lung, kidney, etc.) expressing the S antigen on the membrane. However, the pathway for eliciting the immune response to the S protein coded by mRNA vaccines presents many peculiarities that need to be elucidated.
A common erroneous idea in the theory backing such mRNA vaccines is considering the Spike protein as a simple “foreign antigen” capable of stimulating immune defences, as it occurs for conventional vaccines. Let us consider the documentation provided for the first registration of Moderna’s mRNA-1273 vaccine to the US Federal Drug Administration [
76]. In the presentation illustrating the immunization process [
77], the LNP loaded with the Spike-encoding mRNA would fuse with the plasma membrane of and release the mRNA into antigen presenting cells (APC), which in turn would manufacture the Spike protein and present it on the membrane to CD4+ T helper cells, CD8+ T cytotoxic cells and B cells. According to this theory, the mRNA vaccine: (a) “provides instruction (Spike protein) directly to the immune system” and (b) “efficiently creates specific immune memory in a natural context (in situ)”. This “theoretical” pathway is illustrated in the upper part (A2) of
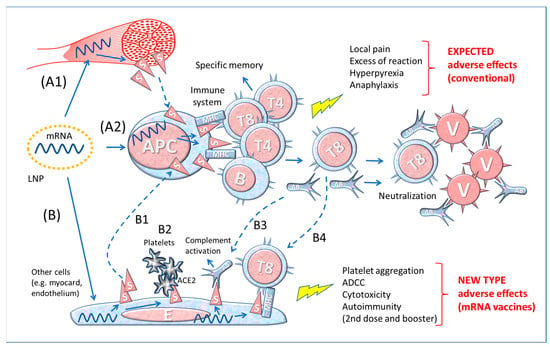
Figure 1.
Figure 1. Diagram of the theory of functioning of anti-COVID-19 mRNA vaccines. A1–2 The simplistic theory representing the production and presentation of Spike protein by APCs to lymphocytes; A1: production of Spike by local muscle cells and release (by shedding) of soluble S that would be captured and processed by APC for immune stimulation; A2: the LNPs transfect the mRNA into APCs, which then produce and present in the context of MHC Spike to immune cells. B. Theoretical consequences of the expression of Spike protein by cell types other than immune cells transfected by mRNA containing LNP. B1: S protein released by somatic cells stimulates the immune system via APC; B2: interaction of blood platelets with S protein on the membrane of endothelial cells; B3: specific antibodies bind to S protein on the membrane of somatic cells (myocardium, endothelium, etc.) and activate the Complement system (or antibody-dependent cytotoxicity; not shown) leading to cell death; B4: specific CD8+ T lymphocytes (T8) attack endothelial cells expressing S protein. Abbreviations and symbols: LNP: lipid nanoparticle; APC: antigen-presenting cell; MHC: Major Histocompatibility Complex; S: Spike; T and B: lymphocytes; V: Virus; E: Endothelial cell; AB: Antibody; ADCC: Antibody-dependent cell cytotoxicity; ACE2: angiotensin-converting enzyme 2. Solid line arrows: action, operation; dashed line arrows: moving, displacement.
This model reproposes the essential steps (not considering the complexities of the MHC system, chemical mediators, and accessory cells, etc.) of the theory of immunization with traditional vaccines made with microbe derivative substances or with the whole microbe after it has been attenuated, inactivated, or killed. However, the mRNA of vaccines is injected into muscular cells which produce and expose on the membrane the Spike protein that eventually could be shed and then captured by the APCs (Figure 1, A1 and A2).
Conventional immunological knowledge teaches that antigen-presenting cells (APCs, dendritic cells, macrophages, and B memory cells) “capture” extracellular potentially pathogenic particles (showing a pathogen-associated molecular pattern, PAMP) by means of a series of appropriate receptors. These antigenic particles are then internalised by endocytosis or phagocytosis (depending on the particle dimension and the cell type), “processed” (i.e., digested) in small peptides (approx. 30 amino acids) and eventually inserted into the MHC-II (major histocompatibility complex) cleft for informing Th (CD4+) lymphocytes. However, for COVID-19 mRNA vaccines the scenario might be not so straightforward, as discussed below.
The theory of “conventional” vaccinology predicts that immunity is obtained by injecting the foreign “antigen”, inactivated so as not to cause any harm to the host, yet still able to stimulate a specific humoral and cellular immune reaction [
78]. According to this view, the expected adverse events following immunization are transient pain and inflammation at the injection site and transitory systemic symptoms such as fever and malaise. Albeit rarely, serious adverse effects may occur after vaccination, for instance due to an allergy condition (anaphylaxis) or to immune dysregulation or autoimmunity mediated by antigens themselves or by the adjuvant (e.g., aluminum particles), or possibly because of inadvertent use of improperly inactivated or mutated microbe (e.g., Sabin polio), or genetic susceptibility [
40,
79,
80,
81,
82].
In the specific case of mRNA-driven antigen delivered via LNP, the following peculiarities should be considered: 1. The LNPs may fuse with the membrane of any cell they encounter and therein release the payload [
83]. This implies that the mRNA may direct the synthesis of the Spike protein not exclusively in muscle cells but also in APCs and other somatic cells. 2. The mRNA is provided with a leader sequence, which directs the synthesis of the Spike protein in endoplasmic reticulum-associated ribosomes. The membrane bound S protein would then travel through the Golgi complex (here it will be split into S1 and S2 by furin) and then be exposed on the plasma membrane via insertional exocytosis [
14]. Transfected cells could free the S protein and/or its fragments following T cell killing, and S1 (which is non-covalently bound to S2) could be shed from the membrane [
14,
22]. Consistently, high levels of soluble Spike proteins are found in the circulation of vaccinees with myocarditis [
84]. The soluble Spike can be subsequently endocytosed by APCs and B lymphocytes. The transfected cells may release exosomes expressing the S protein on the membrane, which also contribute to immunostimulation of APCs in distant organs [
66].
Antigen processing follows two different routes depending on the cell type (immune or non-immune) and whether the antigen locates in the endosomal compartments or in the cytoplasm. In the former case (occurring for instance in APCs), the exogenous antigen internalized via endocytosis/phagocytosis is proteolyzed by the endosomal cathepsins and the fragments inserted into the cleft of the MHC class II antigen (HLA-II) to be exposed on the plasma membrane for informing the CD4+ T helper lymphocytes. In the case of the virus infection (e.g., SARS-CoV-2) of parenchymal cells, viral proteins (for instance the S protein) in the cytoplasm are proteolyzed by the ubiquitin-proteasome pathway and the (immunodominant) peptides translocated into the endoplasmic reticulum where they are inserted into the cleft of MHC class I and eventually exposed on the plasma membrane. This will inform CD8+ T cytotoxic lymphocyte that the cell has been infected and should be killed. B lymphocytes, on their side, are stimulated by soluble antigens recognized by membrane B-cell receptors (a complex containing IgD or IgM) to become plasma cells producing and secreting soluble antibodies. There is crosstalk of cytokines between APCs, Th, Tc and B lymphocytes to orchestrate the immune response.
However, in the case of the COVID-19 vaccination with LNP loaded with the modified mRNA we face unpredicted outcomes, since the mRNA transfection could aspecifically occur in any cell, including APCs, endothelial cells, and parenchymal cells of distant organs, wherein the mRNA would then direct the persistent synthesis of the modified (stabilized in open conformation) S protein. The processing route of the S protein will determine the fate of the transfected cells.
In case of LNP transfection of parenchymal cells (ideally only the muscle cells at the injection site), the exposure on the membrane of the S protein would predictably trigger the CD8+ T lymphocyte cytotoxicity, much like what would happen to virus-infected cells. Yet, at variance from natural infection with SARS-CoV-2, in the transfected cells the S protein may (in part) not be processed, and be exposed on the membrane not in the context of the MHC class I. This eventuality could deceive the immune cells, which could consider the protein as a self.
To add complexity, we must consider that other cells, in addition to APCs, can be transfected by the mRNA containing LNPs, as represented in Figure 1, bottom drawing (B). These cells would produce Spike proteins, display them on the membrane (or release after cell death or shed the S1) and trigger the response of the immune system (B1 in Figure 1). Furthermore, the Spikes exposed on the membrane of endothelium can interact with the ACE2 receptors exposed on the platelet membrane, favouring their aggregation (B2 in Figure 1). When the Spike synthesis is induced by boosters, i.e., in immunized individuals, the risk is that the transfected cells become victims of the aggression by previously formed antibodies (B3 in Figure 1) or by cytotoxic T8 lymphocytes (B4 in Figure 1). If this is the case, the adverse events following repetitive immunizations may be worse and involve various organs in which the Spike localizes.
Hence, the mRNA vaccine “theory” neglects the possibility that any cell producing the Spike protein and displaying it on its membrane (associated or not with MHC-I) will be attacked and destroyed by CD8+T cells. The severity of the consequences for the host following the vaccination will depend on the type and number of cells affected and the tissue where the reaction occurs. For example, myocarditis is considered an adverse reaction to mRNA vaccination [
85,
86]. The facts that this event is more frequent after the second dose and it occurs a few days after the inoculation [
27], suggest an immune-mediated mechanism analogous to an auto-immune reaction. To conclude, the Spike protein acts in a peculiar way, not simply as an immunogen, but as a disease-causing agent.
2.3. Differences between Contact with the Whole Virus and Vaccine-Derived Spike Protein
Contact with the whole virus comprehensively instructs the immune system and all its components, therefore in case a constituent of the virus changes because of gene mutations, the immune memory toward the conserved viral components can still trigger the immune response. Furthermore, the different fragments of the virus presented by APCs to the lymphocytes trigger a complex polyclonal immune response that effectively neutralizes the virus.
The components of a virus shape the type of the innate and specific immune response. A pathogen contains proteins, lipids, carbohydrates, and nucleic acids that constitute the so-called PAMPs that bind to the PRRs (Pattern Recognition Receptors) present on the APCs. The interaction leads to the maturation of the APC and the initiation of the adaptive immune response with the priming and differentiation of the antigen specific T helper cells, T cytotoxic cells, and B cells. The PAMPs combination determines the type (innate and/or adaptive), the extent, and the duration of the immune response. The biological and immunological implications of Spike immunization in relation to the type of vaccine, adjuvant, and route of administration have been studied in animal models [
87].
All viruses have a specific cellular tropism, meaning that they enter and infect only those cells expressing the suitable receptor on their membranes. In the case of SARS-CoV-2, the virus preferentially enters the cells expressing the receptor for Spike (i.e., ACE2). On the contrary, as outlined above, the mRNA vaccines delivered via LNP can in principle (and in practice) transfer the information for the synthesis of the S protein to any cell.
Many things about the vaccination outcome are still unknown: 1. Is the amount of S protein synthesized upon vaccination comparable with that of a natural virus infection or is it higher by many orders of magnitude? 2. How long does the Spike synthesis last following administration of mRNA? 3. How long do vaccine-derived Spike proteins remain biologically active?
It is difficult to calculate exactly the number of copies of the Spike protein that results from the administration of these vaccines, because the declared amount of mRNA is not consistent in all batches (the producer Pfizer admitted that only 30 to 70% of the mRNA in the vaccine is integer for effective translation) and because its intracellular stability may vary from cell to cell.
Thus, it is reasonable to expect a big difference in the biological effect and the immune response between the natural infection and the administration of mRNA vaccines.
This entry is adapted from the peer-reviewed paper 10.3390/biomedicines11020451