Some neurogenic stimuli, such as physical exercise, can partially revert the decrease of neurogenesis occurring during aging [
208,
209,
210], while other neurogenic stimuli, though effective in the adult, are unable to elicit neurogenesis in aged models. Between them are treatment with antidepressant molecules regulating 5-hydroxytryptamine (5-HT; selective 5-HT reuptake inhibitors, i.e., SSRI) or norepinephrine pathways [
211,
212,
213], and learning [
214,
215]. Concerning nutrients, we have recently shown that HTyr activates hippocampal neurogenesis, including stem cells, in aged mice [
47]. The majority of these neurogenic stimuli activates only progenitor cells and not stem cells, for example running [
209] or the antidepressant fluoxetine [
216,
217,
218]. However, some nutrients are also able to activate stem cells, though not in aged mice, but in models that are defective for neurogenesis, as for instance the natural flavonoid luteolin in a mouse model of Down syndrome [
219,
220]. Additionally, HTyr increases the survival of active (c-fos+) new neurons (see
Section 10.3) and decreases the expression of the markers of aging and neuroinflammation lipofuscin and ionized calcium-binding adapter molecule 1 (Iba1) [
47]. Overall, the finding that HTyr reactivates aging stem cells supports the idea of using dietary supplements to counteract cognitive deterioration during aging. Another report demonstrated that HTyr treatment of prenatally stressed rat mothers (by restraint stress on days 14–20 of pregnancy) restores in the offspring of treated mothers the mRNA levels of BDNF, GAP43, synaptophysin, and
N-methyl-D-aspartate (NMDA) receptor subunits NR1, NR2A, and NR2B, all neural markers involved in synaptic plasticity and decreased by stress in the entire hippocampus [
221]. However, no information was given regarding the generation of new neurons in the neurogenic niches [
221]. More recently, a report about treatment with HTyr in vivo after stroke, obtained with transient occlusion of the middle cerebral artery, showed that HTyr-fed mice presented improved short-term recognition memory and an increase of BDNF and of cerebral blood flow in the hippocampus. Moreover, the study suggested a trend of increase of dentate gyrus total progenitor cells (DCX+ cells), without, however, showing a significant difference between treated and untreated conditions after stroke [
222]. This may also depend on the dosage of HTyr treatment, lower than that used in the study by D’Andrea et al. (2020) [
47]. This report, together with that by [
221], brings to evidence the synaptogenic potential of HTyr, an important feature in the process of neural regeneration occurring after stroke. Additionally, OLE treatment, in a rat model of stroke by occlusion of the middle cerebral artery, induces a decrease of cerebral edema and plasma fibrinogen, inhibition of angiotensin converting enzyme (ACE) activity, and an increase of the antioxidant enzymes SOD, GPX, and catalase in brain tissue [
223]. Another previous report in a mouse model of stroke by occlusion of the middle cerebral artery followed by reperfusion indicates that OLE (100 mg/kg i.p.) reduces the volume of cerebral infarction and apoptosis through reduction of Bax and increase of Bcl2 expression [
224]. Likewise, OLE treatment after brain stroke caused in rats by intracerebral hemorrage following a collagenase injection in the brain improves brain edema and protects the integrity of the blood–brain barrier [
225].
HTyr treatment in vivo of rats (10 mg/Kg) also mitigated spinal cord injury after laminectomy, since the neural function was rescued and lipid peroxidation and myeloperoxidase activities were reduced, together with decrease of proinflammatory cytokines and apoptotic markers [
234].
7. HTyr and OLE in Senescence and Lifespan
7.1. Effects of HTyr and OLE on Senescence
Cellular senescence is characterized by a permanent arrest of cell proliferation due to stress conditions, including telomere shortening, oxidative stress, hypoxia, oncogenic activation, and DNA damage, and has been linked to processes such as tumor suppression, tissue repair, embryogenesis, and aging. The principal hallmarks of senescence are an increase in senescence-associated βgalactosidase activity (SA-β Gal), the presence of telomere dysfunction-induced foci (TIF), upregulation of specific cell cycle regulators (mainly the p53-p21 and p16-pRb axes), altered gene expression patterns, and the activation of a senescence-associated secretory phenotype (SASP), implying the synthesis and secretion of inflammatory mediators, growth factors, and extracellular matrix proteins [
270,
271]. The SASP is thought to play a major role in many age-related diseases, such as AD [
272] and cancer [
273], where it contributes to the maintenance of chronic inflammation, and in cardiovascular diseases and type 2 diabetes, also characterized by a low-grade chronic inflammation state.
It is worth noting that the elimination of senescent cells in progeroid and aged mice, through senolytic drugs or genetic mutations, markedly improves health span after senescent cells are cleared [
270,
274].
Several papers have reported that HTyr and OLE are endowed with the ability to modulate senescence or related inflammation in primary mammalian cell cultures, which represent suitable cellular models to study in vitro the mechanisms of aging. In pre-senescent human lung cells (MRC5) and neonatal human dermal fibroblasts (NHDFs), chronic (4–6 weeks) treatment with 1 μM HTyr or 10 μM OLE aglycone reduced some known markers of senescence, such as p16 and SA-β Gal. Moreover, the treatment decreased the secretion of senescence/inflammation markers such as IL-6 and metalloprotease, and levels of COX-2 and α-smooth-actin. Furthermore, the induction of inflammation following exposure to TNFα was abolished in OLE- and HTyr-pre-treated NHDFs [
275]. Thus, the modulation of the senescence and senescence-associated inflammatory phenotype might be an important mechanism underlying the beneficial effects of olive oil phenols. Moreover, OLE (5 μM) and HTyr (1 μM) exerted a protective effect on 8 Gy irradiation-induced senescence, in γ-irradiated NHDFs, mitigating DNA damage and reducing the SASP mediated by the cyclic GMP AMP synthase/stimulator of intereferone genes (cGAS/STING)/NFκB pathway [
276]. The ability of OLE and HTyr to alleviate senescence status, and the related SASP in normal cells can be exploited to improve the efficacy and safety of cancer radiotherapy.
Similar results were obtained in human dermal fibroblasts (HDFs) exposed to UVA. In this cell line, HTyr decreased SA-β-galactosidase activity and expression levels of MMP-1 and MMP-3 in a dose-dependent manner. These observations were accompanied by an anti-inflammatory effect, demonstrated by the reduced expression of IL-1β, IL-6, and IL-8 genes [
277]. Interestingly, Varela-Eirín et al. (2020) demonstrated that OLE has senolytic activity in osteoarthritic chondrocytes (OACs) [
278]. In fact, OLE reduced the inflammatory and catabolic factors mediated by NF-kB (IL-1ß, IL-6, COX-2, and MMP-3) and lowered cellular senescence in OACs, synovial and bone cells.
7.2. Effects of HTyr and OLE on Lifespan
Proliferative cellular lifespan is a complex process that is governed by multiple pathways in part interdependent with cellular senescence. In fact, normal somatic cells in vitro, after a finite number of cell divisions, can enter in the non-proliferative state of senescence [
279].
In in vitro models, it was observed that treatment with OLE extended the lifespan of normal human fibroblasts (NHFs) by enhancing the activity of proteasome [
280], which is known to decline during aging in human tissues as well as in senescent primary culture cells. Moreover, HTyr treatment was able to increase chronological lifespan (CLS) in quiescent NHFs, which have lost their capacity to repopulate. CLS measures the length of time nondividing cells survive before reach senescence, without telomere attrition. This process could be related to the observation that quiescent human somatic cells exhibit age-dependent loss in their regenerative capacity. The observed HTyr-induced extension of CLS was associated with an increase in manganese superoxide dismutase (SOD2) activity and reduction of age-associated mitochondrial ROS generation [
281].
It is noteworthy that, as mentioned in previous sections, HTyr and OLE have shown the ability to activate AMPK and SIRT1 pathways and inhibit the mTOR and insulin/IGF1 signaling, which are crucially involved in regulation of health span and, as a consequence, lifespan and senescence in animal models. Interestingly, numerous studies have been focused on the effects of HTyr and OLE on the lifespan of
Caenorhabditis elegans. This free-living nematode is a powerful animal model mostly used in aging research due to its short life cycle. It was recently observed that OLE could significantly prolong the lifespan of
C. elegans increasing the survival rates of worms against lethal heat shock and oxidative stress. OLE supplementation increased the expression and activity of antioxidant enzymes and suppressed the generation of malondialdehyde in nematodes. These positive effects on longevity and stress resistance were mediated by the factors DAF-16/FoxO, which plays a vital role in the insulin/IGF-1 signalling (IIS) pathway, and the SKN-1/Nrf2 pathway, another stress response and longevity pathway [
282].
HTyr was also able to increase the survival of worms after heat stress and could further prolong the lifespan in unstressed conditions. In addition, in aged worms, exposure to HTyr and OLE led to the improvement of locomotive behavior and the attenuation of autofluorescence as a marker of ageing [
154].
Moreover, numerous studies have been conducted on the beneficial effects of olive oil polyphenols in
C. elegans models of PD and AD. Although
C. elegans is not able to develop PD, exposure to the pesticide rotenone or the transgenic expression of human α-synuclein induces the Parkinsonian-like syndrome in
C. elegans, which manifests as impaired movement. In models of PD, HTyr and OLE, as already mentioned in
Section 6.2, have been shown to enhance swim performance of worms, to reduce α-synuclein accumulation in muscle cells, and to prevent neurodegeneration in α-synuclein-containing dopaminergic neurons [
154]. Analogously, the treatment with a natural extract enriched in HTyr showed beneficial effects on the lifespan of wild-type nematodes and in a PD-model [
155]. More recently, it was observed that in
C. elegans, treatment with an olive fruit extract 20% rich in HTyr had beneficial effects on AD features such as Aβ- and tau-induced toxicity, as well as on oxidative stress. The treatment prevented oxidative stress and delayed Aβ-induced paralysis reducing Aβ aggregates. Indeed, the extract partially reduced the proteotoxicity associated with aggregation of the tau protein. The observed effects were correlated to the activity of the SKN-1/NRF2 transcription factor and the overexpression of HSP-16 [
283].
8. Protective Effects of HTyr and OLE against Skeletal Muscle Dysfunction
In addition to its fundamental function in locomotion and maintenance of posture, skeletal muscle plays crucial roles in energy and glucose metabolism. This tissue has also emerged as an endocrine organ producing and releasing growth factors and cytokines (commonly termed myokines) that modulate systemic physiology. Furthermore, skeletal muscle may cross-talk with the nervous system and other tissues via direct muscle–nerve interactions, release of metabolites, systemic adaptation deriving from the energy demand of contracting muscle (e.g., during exercise), and myokines [
21,
284]. The age-associated loss of muscle mass and strength (called sarcopenia) is an important medical condition that results in fragility and reduced mobility and negatively affects bone mass, which also declines during aging causing osteopenia and osteoporosis. The concomitance of sarcopenia and osteoporosis in elderly patients leads to severe physical disability, frailty, high risk of falls, and osteoporotic fractures [
285]. Furthermore, sarcopenia also influences global metabolic homeostasis, lifespan, systemic aging, and progression of age-related degenerative diseases in non-muscle tissue, as well as the organism’s responses to oxidative stress and dietary restriction [
286].
8.1. Age-Related Sarcopenia and Osteoporosis
Multiple factors contribute to sarcopenia of aging, including chronic activation of inflammatory pathways, enhanced ROS generation and/or dysregulation of redox signaling, loss of muscle proteins resulting from an imbalance between protein synthesis and degradation, dysregulation of muscle autophagy, and impairment of neuromuscular transmission. These age-associated dysfunctions lead to myofiber death, muscle atrophy, and reduced regenerative potential [
287,
288].
A few studies have shown beneficial effects exerted on sarcopenia by olive mill wastewaters- and olive leaf-derived extracts rich in HTyr and/or OLE.
Pierno and colleagues (2014) tested the therapeutic potential of a polyphenolic mixture (LACHI MIX HT), extracted from olive mill wastewaters and containing mainly HTyr and low amounts of other phenolic compounds (tyrosol, catechol, gallic acid, homovanillic acid, and caffeic acid) in the amelioration of skeletal muscle dysfunctions due to aging-associated oxidative stress [
289]. The effects of treatment of old rats with LACHI MIX HT were compared with those of purified HTyr. LACHI MIX HT treatment was able to counteract some of the alterations in excitation/contraction function typically observed in aged skeletal muscle. In particular, the treatment was shown to increase muscle weight, to improve sarcolemma chloride channel conductance, contractile function, and ATP-dependent potassium channel activity and to decrease blood creatine kinase levels and the resting cytosolic calcium concentration. Furthermore, LACHI MIX was more effective than purified HTyr in ameliorating most of the examined parameters, possibly due to the synergic activity of its various components.
Extracts from olive tree (
Olea europaea) leaves are rich in OLE and HTyr, have a high antioxidant and scavenging power, and have been reported to exert beneficial effects on several health conditions, including hypertension, cardiovascular diseases, diabetes, and hyperlipidemia [
290]. A recent study analyzed the effects of treatment with an olive leaf extract on aging-related sarcopenia and skeletal muscle insulin resistance [
291]. Sarcopenia is known to be associated with: (i) increased expression of the E3 ubiquitin ligases Muscle Ring-finger protein-1 (MURF1) and Atrogin-1, which play a major role in the process of age-induced muscle loss [
292]; (ii) increased expression of myostatin, a secreted myokine that inhibits skeletal muscle growth and negatively impacts muscle metabolism [
293,
294]; (iii) upregulation of histone deacetylase-4 (HDAC4), a deacetylase that promotes muscle atrophy in response to multiple stimuli by targeting several substrates, including myosin heavy chains (MyHC) and the transcriptional regulator PGC1α [
295]; (iv) upregulation of the myogenic regulatory factors MyoD and myogenin, possibly as a compensatory mechanism [
296]. A 21-day treatment of old rats with an olive leaf extract was shown to attenuate the age-induced atrophy of gastrocnemius muscle and the alterations in the mRNA expression of many of the above- mentioned markers of sarcopenia. Specifically, the authors observed a reduction of HDAC4, MyoD, myogenin, myostatin levels, and upregulation of PGC1α levels. The treatment also prevented the age-induced elevation of several pro-inflammatory markers and enhanced the expression of the anti-inflammatory cytokine IL-10 that was reduced in old untreated animals. Furthermore, the treatment improved muscle insulin sensitivity in gastrocnemius muscle by activating the insulin-dependent PI3K/Akt pathway [
291].
In a follow-up study, the same authors evaluated whether the addition of the olive leaf extract to an oil mixture, composed of 25% algae oil and 75% EVOO (AO:EVOO), could synergistically attenuate sarcopenia in old rats [
297]. In previous studies, the AO:EVOO mixture per se was reported to exert protective effects against cardiometabolic and skeletal muscle alterations associated with aging [
298,
299]. Combined treatment of 24-month-old rats with the olive leaf extract and AO:EVOO mixture provided further benefit to sarcopenia preventing not only the decrease in the weight of gastrocnemius muscle but also in the weight of the soleus muscle. Besides preventing the induction of the sarcopenia-related markers HDAC4, MyoD, myogenin, and rescuing the downregulation of PGC1α, the co-administration of both ingredients attenuated the reduction of MyHC isoforms, downregulated the expression of Atrogin-1 and MuRF1, prevented age-induced macrophage infiltration, and activated the insulin-dependent PI3K/Akt pathway in gastrocnemius muscle and adipose tissue [
297].
There is ample evidence for a constant cross-talking between skeletal muscle and bone via paracrine and endocrine signals and for the existence of common pathogenic pathways shared by sarcopenia and osteoporosis including oxidative stress, increased inflammatory cytokine production, reduced anabolic hormone secretion, and reduced physical activity [
300,
301]. Consumption of olives, olive oil and olive polyphenols has been shown to improve bone health [
302,
303]. In particular, a number of studies reported physiological effects exerted by HTyr and/or OLE against osteoporosis. Hagiwara et al. (2001) evaluated the effects of HTyr and OLE on bone formation using cultured osteoblasts and osteoclasts, and on bone loss in ovariectomized mice, a widely used experimental model of post-menopausal osteoporosis [
304]. They showed that HTyr and OLE inhibited the formation of multinucleated osteoclasts in culture, and that OLE enhanced the deposition of calcium by osteoblasts. Furthermore, OLE and HTyr were able to decrease bone loss in trabecular bone of femurs in ovariectomized mice. Other studies reported that OLE, HTyr, and olive mill wastewater extracts elicited beneficial effects on femur bone mineral density and improved inflammatory and oxidative stress markers in an experimental model of senile osteoporosis induced by ovariectomy associated with an acute inflammation in rats [
305,
306]. More recently, a beneficial effect of OLE on bone mineral density of lumbar vertebra and left femur of ovariectomized rats was also reported by Liu et al. (2022) [
307]. Moreover, OLE could reduce the serum levels of IL-6 and malondialdeyde (a marker of lipid oxidation). In cellular experiments, it could also promote the proliferation of primary osteoblasts while inhibiting osteoclast differentiation by upregulating expression of osteoclastogenesis inhibitory factor (OPG) and downregulating expression of receptor activator for nuclear factor-
κB ligand (RANKL). Finally, a study conducted in the murine osteoblast MC3T3-E1 cell line revealed a cytoprotective effect of HTyr against oxidative stress-induced osteoblast apoptosis and shed light on the underlying mechanism [
308]. In fact, HTyr was shown to prevent oxidative stress-induced mitochondrial dysfunction and osteoblast apoptosis by decreasing the cleavage of optic atrophy 1 (OPA1), one of the most important mitochondrial dynamics proteins, and by activating the Akt/glycogen synthase kinase 3 β (GSK3β) signaling pathway.
8.2. Skeletal Muscle Atrophy and Oxidative Stress
In addition to aging, skeletal muscle atrophy is the consequence of various conditions, such as muscle disuse, strenuous exercise, denervation, neurodegenerative disease, muscular dystrophies, obesity, and diabetes.
Periods of immobilization are often associated with aging. Prolonged periods of muscle inactivity result in oxidative stress and chronic elevation of ROS production, primarily derived from mitochondria, within inactive muscle fibers. Such disturbed redox signaling critically contributes to disuse-induced muscle atrophy through several mechanisms, including (i) inhibition of muscle protein synthesis via repression of the anabolic Akt/mTOR signaling; (ii) activation of the Ca
2+-activated proteases calpains; (iii) acceleration of protein degradation via the ubiquitin–proteasome system; (iv) activation of apoptosis; (v) stimulation of autophagy [
309]. By using a rat model of unloading-induced muscle atrophy (3-week tail suspension), Liu et al. (2014) showed that 7 days’ reloading efficiently rescued skeletal muscle atrophy and mitochondrial dysfunctions [
310]. Interestingly, administration of a mixture of mitochondrial nutrients including HTyr, α-lipoic acid, acetyl-L-carnitine, and coenzyme Q
10 for 4 weeks to unloaded rats exerted a reloading-like effect and promoted the recovery initiated by reloading. Specifically, treated animals showed enhanced motor function, increased soleus muscle weight, decreased protein degradation, and apoptosis. Furthermore, the nutrient mixture rescued unloading-induced mitochondrial defects, by increasing total anti-oxidative capability, mitochondrial copy number and electron transport chain complex I and II activities and markers of mitochondrial biogenesis, such as PGC1α, nuclear respiratory factor 1 (NRF1), and mitochondrial transcription factor A (Tfam).
Redox-sensitive signaling pathways also play a pivotal role in exercise-mediated remodeling of skeletal muscle. Skeletal muscle shows different responses to the type of exercise, as well as its frequency, intensity, and duration. Increased ROS production in contracting muscle fibers plays a required role in skeletal muscle adaptations induced by regular endurance or resistance exercise training, while excessive ROS produced by strenuous or acute exercise can cause muscle oxidative stress, fatigue, and damage [
309,
311,
312]. As reported by Feng et al. (2011), an 8-week intensive exercise program in rats resulted in decreased endurance capacity and highly induced expression of muscle atrophy (MURF1 and Atrogin-1) and autophagy markers (Atg7, Beclin-1 and LC3) [
313]. This was accompanied by increased mitochondrial fission induced by excess of ROS in skeletal muscle and a decrease in PGC1α and complex I subunit expression. All of these changes were eliminated by treatment of exercised rats with HTyr. In addition, HTyr enhanced mitochondrial fusion and mitochondrial complex I and II activities and inhibited the expression of the oxidative-stress-responsive proteins p53, p21, and SOD2 in muscles of exercised rats [
313]. Therefore, HTyr appears to improve exercise capacity by protecting contracting muscles from excessive ROS production.
8.3. Oxidative Damage and Skeletal Muscle Cell Degeneration
Protective effects of HTyr and HTyr derivatives on oxidative stress- and inflammation-induced mitochondrial dysfunction and muscle cell degeneration were also described in the C2C12 myogenic cell line.
Wang et al. (2014) reported that induction of oxidative stress in C2C12-differentiated myotubes promoted significant mitochondrial dysfunction in a time-dependent manner, accompanied by decreased expression of MyHC and myogenic regulatory factors (MyoD, myogenin, MRF4) and induction of apoptosis [
314]. Furthermore, oxidative stress was shown to promote rapid cleavage of OPA1, a protein that controls mitochondrial inner membrane fusion and remodeling. Pre-treatment with HTyr acetate, which is present in olive oil at a concentration similar to that of HTyr, significantly prevented oxidative stress-induced OPA1 cleavage and mitochondrial morphology changes, and this was accompanied by improvement of mitochondrial oxygen consumption capacity, ATP productive potential and activities of mitochondrial complexes I, II, and V, and inhibition of oxidative stress-induced MyHC decrease [
314].
Another study showed that pre-treatment with HTyr or the corresponding ester HTyr laurate effectively protected C2C12 myoblasts from apoptotic cell death induced by H
2O
2 treatment [
105].
Furthermore, pre-treatment with HTyr was shown to counteract muscle cell degeneration induced by the inflammatory cytokine TNFα in C2C12 differentiating myoblasts by increasing the expression of differentiation markers (MyHC and myogenin), PGC1α and mitochondrial complexes I and II, as well as the activity of muscle creatine kinase [
315].
Finally, a couple of studies investigated the effects of OLE in primary avian skeletal muscle cells. Kikusato and colleagues (2016) reported that treatment of avian muscle cells with OLE induced the mRNA expression of avian-specific uncoupling protein (avUCP), PGC1α, and downstream mitochondrial biogenesis-related genes (NRF1, Tfam, ATP5a1), as well as the activity of mitochondrial cytochrome c oxidase (COX) [
316]. Additionally, the expression of the SIRT1 deacetylase was found upregulated by OLE. Since it is well-known that deacetylation by SIRT1 induces the PGC1α co-transcriptional activity [
181], these results suggested that OLE upregulates mitochondrial biogenesis and avUCP expression in muscle avian cells through the SIRT1/PGC1α regulatory axis. Furthermore, OLE was observed to suppress the levels of mitochondrial ROS generation, possibly via the up-regulation of SOD2 and avUCP gene expression. In a follow-up study, Muroi et al. (2022) used a specific inhibitor of SIRT1 activity to confirm that indeed it can mediate the enhancement of PGC1α, avUCP, ATP5a1, and SOD2 gene expression and the suppression of mitochondrial ROS generation induced by OLE in chicken muscle cells [
317]. The study also showed that the action of OLE on the above markers of mitochondrial biogenesis and oxidative damage can be mediated by transient receptor potential cation channel subfamily V member 1 (TRPV1), as determined by using an antagonist of TRPV1. Furthermore, treatment of chicken muscle cells with OLE was shown to increase intracellular Ca
2+ concentration and stimulate the phosphorylation, and hence activation, of the AMPK kinase in a TRPV1-dependent manner. Based on the knowledge that Ca
2+/calmodulin-dependent protein kinase 2 induces phosphorylation of AMPK and that AMPK, SIRT1, and PGC1α are part of a highly coordinated regulatory network [
181,
318], the authors proposed that the effects of OLE in chicken muscle cells may be due to induction of Ca
2+ influx, possibly through the activation of TRPV1 localized at ER, and increased activity of AMPK, followed by SIRT1 activation and then activation of PGC1α and induction of downstream mitochondrial targets [
317].
8.4. Skeletal Muscle Insulin Resistance and Metabolic Syndrome
As mentioned earlier, skeletal muscle plays a crucial role in whole-body energy homeostasis and is a primary tissue of insulin-induced glucose uptake and oxidative metabolism. Muscle atrophy and sarcopenia are associated with the development of skeletal muscle insulin resistance [
319], which is considered to be a major driver of metabolic syndrome (MetS), a cluster of medical conditions (obesity, hypertension, dyslipidemia, hyperglycemia) that together increase the risk of developing type 2 diabetes, cardiovascular diseases, and stroke. Major factors in the development of MetS include sarcopenia, chronic inflammation, abdominal obesity, insulin resistance/hyperinsulinemia, physical inactivity, high fat intake, and genetic factors. MetS, obesity, and diabetes induce adverse effects on skeletal muscle function, including muscle fiber atrophy and contractile dysfunctions, altered metabolism, insulin resistance, oxidative stress, mitochondrial dysfunctions, and reduced regenerative potential [
319,
320,
321,
322,
323].
Commonly used MetS murine models exhibit metabolic disorders induced by high-fat diet (HFD) feeding or a non-functional leptin pathway, such as the
db/
db mouse [
328]. Cao and colleagues extensively examined the effects of HTyr in skeletal muscle and liver of C57BL/6 mice fed a HFD with or without supplementation of low-dose or high-dose HTyr (10 or 50 mg/Kg/day, respectively) for 17 weeks [
329]. HTyr administration was shown to effectively inhibit body and organ weight increase (50 mg/Kg/day). Both low- and high-dose HTyr treatment enhanced glucose tolerance and lowered the serum levels of glucose, insulin, lipids, and inflammatory cytokines. In skeletal muscle and liver, HTyr reduced the accumulation of lipid deposits through inhibition of the sterol regulatory element-binding transcription factor (SREBP) pathway, attenuated oxidative stress by enhancing antioxidant enzyme activity, elevated the decreased expression of complex I and II subunits of the electron transport chain induced by HFD, and lowered the expression of markers of mitochondrial fission and apoptosis. Moreover, in muscle tissue, HTyr decreased the level of mitochondrial protein carbonylation and elevated the activities of complexes I, II, and IV. The effects of HTyr were also examined in diabetic
db/
db mice treated for 8 weeks with low-dose HTyr (10 mg/Kg/day) or with metformin, a commercial antidiabetic drug, as a positive control. Both HTyr and metformin reduced the fasting blood glucose levels in
db/
db mice, whereas the fasting serum levels of lipids (triglyceride, cholesterol) were lowered by HTyr but not by metformin. Furthermore, HTyr was shown to be more effective than metformin in reducing protein and lipid damage in skeletal muscle and liver and in increasing the activities of mitochondrial complexes I and IV [
329].
Another study evaluated the effects of HTyr acetate (HTyr-Ac) on glucose consumption in C2C12 skeletal muscle cells and 3T3-L1 adipocytes. Treatment with increasing concentration of HTyr-Ac was reported to stimulate glucose uptake in a dose-dependent manner in both C2C12 myotubes and differentiated adipocytes. Furthermore, HTyr-Ac was shown to exert an anti-adipogenic effect by inhibiting adipocyte differentiation and by stimulating lipolysis of fully differentiated adipocytes [
330].
With regard to OLE, it has been shown that C57BL/6 mice fed an HFD supplemented with 0.038% of OLE, for 12 weeks, exhibited reduced levels of fasting glucose and improved insulin resistance. Moreover, the gastrocnemius muscle of OLE-fed mice displayed increased expression and membrane localization of glucose transporter type 4 (GLUT4) protein [
331]. In the same study, C2C12 myotube cells treated with OLE displayed enhanced glucose uptake and GLUT4 translocation to the plasma membrane. Furthermore, OLE improved insulin sensitivity in C2C12 cells treated with palmitic acid, a model of lipotoxicity and insulin resistance. OLE did not show a synergistic effect with insulin regarding glucose uptake, and stimulated phosphorylation, and hence activation, of AMPK but not phosphorylation of the Akt kinase, which is downstream of the insulin signal [
331]. Therefore, the mechanism through which OLE can induce glucose uptake in muscle cells appears to be linked to the activation of AMPK but does not require activation of the PI3K/Akt pathway.
Similar results were reported by Hadrich et al. (2016) who showed that treatment of C2C12 myotubes with OLE can stimulate glucose uptake and activate AMPK and MAPK signaling but not the PI3K/Akt insulin signaling pathway [
332]. In addition, the treatment was shown to protect C2C12 myotubes against oxidative stress induced by H
2O
2 by decreasing ROS production and lipid peroxidation levels.
In agreement with the previously discussed studies, Alkhateeb et al. (2022) have recently reported protective effects of OLE on muscular insulin resistance in isolated soleus muscle preparations treated for 12 h with a high concentration of palmitate [
333]. In fact, OLE treatment enhanced insulin-stimulated glucose uptake, the translocation of GLUT4 at the plasma membrane, and the levels of Akt substrate of 160 kDa (AS160) phosphorylation. Furthermore, OLE promoted the activation of AMPK and an inhibitor of AMPK blocked OLE-stimulated glucose uptake, GLUT4 translocation, and AS160 phosphorylation. This indicated that OLE can ameliorate palmitate-induced insulin resistance via an AMPK-dependent mechanism.
As mentioned earlier, OLE is the most abundant phenolic compound in extracts from olive leaves. By using a rat model of STZ-induced diabetes, Giacometti et al. (2020) investigated the effects of an OLE-rich olive leaf extract on GLUT4 expression and intracellular vesicular GLUT4 trafficking in soleus muscle [
334]. Diabetic rats treated for 10 days with 512, 768 or 1024 mg/Kg of olive leaf extract (containing 20.3, 33 or 44.5 mg/kg of OLE, respectively) displayed significantly reduced blood levels of glucose and triglycerides. Histopathological examination of the diabetic soleus muscle revealed a decrease in fiber size and increased fibrosis, and such pathological muscle alterations were improved after treatment with the olive leaf extract in a dose-dependent manner. Furthermore, treatment with olive leaf extract was shown to promote GLUT4 translocation to the myofiber membrane in soleus muscle and to enhance the expression levels, and the colocalization with GLUT4, of the Rab GPTases Rab8A, Rab13, and Rab14, which are central regulators of vesicular transport along exocytic, endocytic, and recycling pathways [
334,
335].
9. Effect of HTyr and OLE in Gut Microbiota–Brain Axis
9.1. Gut Microbiota
Humans are defined as super-organisms or holobionts that live in harmony with their symbiotic roommates represented by more than 100 trillion microorganisms whose coordinated actions are thought to be important to human life [
336]. Altogether they form anatomical, physiological, immunological, or evolutionary units. In particular, the human gut harbors a bacterial ecosystem of 10
13–10
14 bacterial cells and it is also populated by viruses, fungi, and protozoa that collectively form a complex microbial community known as the gut microbiota [
337]. All these microorganisms are not identified as pathogens by our immune system but, on the contrary, most of them coexist symbiotically with the enterocytes [
338].
The presence of an enormous number of microbes, which far exceeds the number of human cells, leads to the assumption that the microbiota can influence the physiology of the host organism. This is even more reasonable if we consider that the microbial genome (microbiome) vastly exceeds the human host genome’s size [
339]. This element alone is sufficient to realize that our symbiotic bacteria are essential for numerous physiological processes that guarantee well-being by ensuring our body’s homeostasis. An imbalanced composition of bacterial populations can be harmful to human health, contributing to the onset of numerous pathologies. However, although most alterations of the gut microbiota turn into an outbreak of diseases and pathologies, actually, a modification in the composition of the gut microbiota is a physiological event. It has been reported that the gut microbiota plays critical roles in the maintenance of human health: (i) taking part in the digestion of food substances, facilitating access to nutrients that would otherwise be inaccessible to the host; (ii) promoting host cells differentiation to protect them from pathogens; (iii) stimulating and modulating the immune system [
340].
The human intestine is composed of a balanced microbiota with two dominant phyla accounting for about 90% of the total, i.e., Bacteroidetes and Firmicutes, and four less represented phyla, such as Proteobacteria, Actinobacteria, Fusobacteria, and Verrucomicrobia [
341]. Such microorganisms can be autochthonous (indigenous) or allochthonous (transient) and are symbionts in most cases. Despite this, they are considered pathogenic when assuming opportunistic behavior to the detriment of the host [
337]. The Firmicutes/Bacteroidetes ratio is an important parameter for emphasizing a potential gut microbiota disorder [
342,
343,
344,
345] but the abundance, diversity, and homogeneity of the intestinal microbiota are also indicators of a state of health. Nevertheless, this ratio can also be linked to physiological changes in bacterial profiles during different stages of life. Indeed, it is low in the first years of life, then increases in adulthood and decreases again in old age [
346]. Overall, the composition of the microbiota is unique to each individual because it can be influenced by different factors acting throughout life [
347]. Indeed, there are many factors such as genetics, diet, environment, exposure to drugs or more generally lifestyle, which influence the composition of the microbiota allowing the proliferation of certain species, rather than others. This aspect is extremely important as the conditions that favor the balanced assembly of microbial populations beneficial to the host determine the establishment of homeostasis defined as “eubiosis”. If, on the other hand, factors come into play that destabilize this condition of equilibrium, the so-called state of “dysbiosis” occurs. Changing the composition and function of the gut microbiota can alter intestinal permeability, digestion, and metabolism as well as immune responses. In particular, an altered state of the gut microbiota causes a pro-inflammatory state, and this condition can lead to the onset of many diseases ranging from gastrointestinal and metabolic conditions to immunological and neuropsychiatric diseases [
348].
9.2. Gut Microbiota across the Lifespan
The intestinal microbiota establishes a co-evolution relationship with the host (which in fact represents its ecosystem) and its development is regulated by a complex interaction between the host and environmental factors, such as diet and lifestyle. For this reason, knowing the transformation of the gut microbiota from birth to old age may shed light on the variation of this community during lifespan and on the possible associations with disease risks. The symbiosis with the microbiota is established from birth and is rewired several times in the first years of life, a period during which children undergo rapid and irreversible growth, showing significant increases in height and weight, and their organs and cognitive abilities undergo great changes [
349]. Numerous scientific papers reported that the establishment of this symbiosis in childhood is of the utmost importance and that imbalances in intestinal microbiota composition during infancy are associated with various metabolic, immune, and neurological diseases. Thus, early childhood offers a unique opportunity to modulate the gut microbiota in order to promote long-term health [
349]. At first, the intestinal microbial flora of the newborn shows a low biodiversity, which however will increase during development. During the first days of life, Proteobacteria and Firmicutes represent the two most abundant phyla in vaginally born infants; while from the 7th to the 15th day after birth, Actinobacteria appeared in the feces of cesarean-delivered infants [
350]. Around the age of three, the intestinal microbiota will move from a highly unstable and poorly differentiated composition to a more stable composition with the typical characteristics of the adult intestinal microbiota [
351]. From this point on, the gut microbiota will rest in a stable state from the third to the seventh decade of life, although the proportions of
Bifidobacteria,
Firmicutes, and
Fecalibacterium prausnitzii tend to decrease with an increase in
Escherichia coli,
Proteobacteria, and
Staphylococcus [
338,
352]. Research carried out on the dynamics of the microbiota in elderly individuals has also provided further information on the possible trajectories of the intestinal microflora throughout human life [
353]. While the composition of the adult human gut microbiota is generally stable if unperturbed, its stability deteriorates in old age [
352] and alterations causing dysbiosis are becoming more and more frequent due to age-related factors. For example, a decrease in
Bifidobacterium and an increase in
Clostridium and
Proteobacteria have been observed in older people [
338]. Given the role of
Bifidobacterium in stimulating the immune system and metabolic processes, its decrease could partly explain the compromised immune system in the elderly [
341]. Furthermore, the greater possibility of developing dysbiosis in old age is a factor to be correlated to the onset of neurodegenerative diseases.
9.3. Gut Microbiota–Brain Axis
The trade-off between gut microbiota and brain is now regarded as a pivotal hub for healthy life and aging. The so-called microbiota–gut–brain axis is considered a neuroendocrine system and implies a bidirectional communication between the gut microbiota and the brain, whose dysregulation has emerged to affect host health and disease [
354]. A healthy host–microbial balance is fundamental to maintain the physical and mental health of both young and elderly populations, while dysbiosis is steadily more implicated in the onset of metabolic, inflammatory, and neurological disorders [
347]. It is now clear that alterations in top-down (brain to gut) communications are associated with gut inflammation syndromes and appetite disturbances, whereas dysregulations in the bottom-up (gut to brain) interactions are associated with alterations in nervous system functions and neurologic pathologies [
355]. The relationship between microbiota and brain is known to be regulated at multiple levels, such as immunological (cytokines), endocrine (cortisol), and neuronal, including both central (CNS) and enteric (ENS) nervous systems interconnected by the vagus nerve [
354].
The short-chain fatty acids (SCFAs), formed by microbial processing of dietary indigestible fibers, are emerging as key players in neuro-immunoendocrine regulation. Indeed, SCFAs can be used locally by colonocytes as an energy source but are also effective in the maintenance of host intestinal barrier integrity and immunity, suppressing cytokines production by myeloid cells and inducing regulatory T-cell differentiation [
356]. At the colon level, SCFAs also induce the secretion of anorexigenic peptides, which act on hypothalamic centers regulating nutritional habits and energy balance [
357]. In addition to local effects, SCFAs can enter the bloodstream to be distributed to other organs and, after crossing the blood–brain barrier (BBB), contribute to its integrity, inducing the expression of tight junction proteins, and can modulate brain and behavior [
358]). Moreover, in the CNS, SCFAs influence neuroinflammation by regulating the maturation and function of microglia as well as by modulating the levels of neurotrophic factors, increasing neurogenesis, contributing to the biosynthesis of serotonin, and improving neuronal homeostasis and function [
356].
The gut microbiota may influence the functions of ENS and CNS nervous systems also by producing metabolites and neurotransmitters with neuromodulatory properties, such as gamma-aminobutyric acid (GABA), noradrenaline, dopamine, serotonin (5-HT), and their precursors (e.g., tryptophan and tyrosine). In turn, the host nervous system modulates the motility of the gastrointestinal (GI) tract and the intestinal barrier homeostasis, sustaining the microbial community [
355].
Studies usin g germ-free (GF) mice (i.e., axenic, free of all microorganisms) or antibiotic-treated specific pathogen-free (SPF) mice (i.e., free of a specific list of pathogens) have provided the strongest proof of the importance of microbiota in gut–brain signaling. GF mice have been demonstrated to have altered behavior, impaired immune systems, dysregulated hormone signaling, abnormal metabolism and neurotransmission with respect to their standard counterparts [
359]. For example, GF mice were reported to show hyperactivity of the hypothalamic–pituitary–adrenal (HPA) axis upon restraint stress, which could be reversed by administration of
Bifidobacterium infantis, but not by monocolonization with the enteropathogenic bacteria
E. coli [
360,
361], highlighting the influence of microbiota on stress responsivity. In addition, a complete SPF flora was able to partly reverse the HPA response to stress only when it was introduced at an early stage of development, suggesting that brain sensitivity to gut signals may occur only within a critical time window [
360]. Anxiety-like behaviors were also found to be affected in GF mice, which frequently showed an increased exploratory and locomotor behavior, as an index of reduced anxiety [
362,
363,
364]. Furthermore, altered expression of synaptic plasticity-related genes, including BDNF and nerve growth factor-inducible clone A (NGFI-A) [
364,
365], as well as variable plasma levels of neurotransmitters [
363], have been reported in GF mice. However, it is worth noting that the results of the studies on GF mice may vary depending on age, gender, and strains used, as reviewed by Cryan et al. (2019) [
347].
Besides being an important risk factor for the development of many diseases, ranging from cardiovascular complications to neurologic illnesses, gut dysbiosis has been also demonstrated to affect post-disease recovery. For example, emerging experimental and clinical evidence showed the influence of gut microbiota not only on ischemic stroke pathogenesis but also on treatment outcomes. In fact, ischemic brain dysregulates intestinal homeostasis directing aberrated signals to the intestine either via the neural or HPA axis pathways, resulting in poor stroke treatment prognosis [
366]. On the other hand, the transplantation of gut microbiota from normal mice into the intestinal tract of mice with ischemic stroke improved the long-term prognosis and survival rate [
367]. Therefore, therapeutic approaches targeting gut dysbiosis can be considered as promising tools for the treatment and management of stroke or, more in general, of various age-related disorders in which gut dysbiosis may have a role, including neurodegenerative, cardiovascular, metabolic, and musculoskeletal diseases, as well as immune system diseases and cancer [
368,
369,
370,
371].
9.4. Effects of HTyr and OLE on Gut Microbiota under Pathological Conditions
It is now well-established that dietary habits represent one of the main factors determining the composition of the gut microbiota. The Mediterranean diet, which is rich in foods of plant origin, is universally recognized as healthy, and it is amply demonstrated that EVOO, its main source of fat, brings benefits to human health influencing the gut microbiota composition [
372]. In general, ingested polyphenols have been reported to counteract oxidative stress and inflammatory injury, thus protecting the digestive system from chronic and recurrent diseases. Furthermore, it is widely recognized that they are able to exert neuroprotection and promote cognitive functions, but their connection with the gut microbiota is a more recent discovery. Several studies reported the beneficial effect of EVOO or, more in general of polyphenols, on intestinal microbiota [
72,
372,
373,
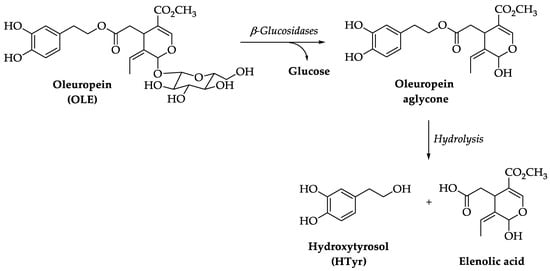
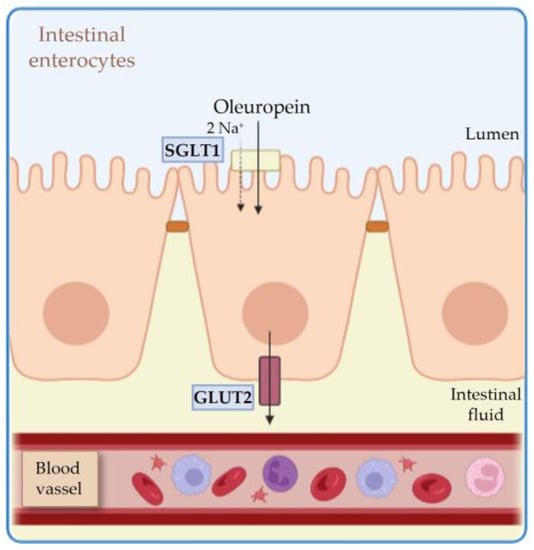
374], whereas less is known about the effect of the exclusive administration of HTyr and OLE. Thus, in this review, we will focus on the outcome of HTyr and OLE on gut microbiota shaping and its implications on healthy host aging. Unlike HTyr and tyrosol, which are well absorbed in the small intestine, OLE reaches the colon unchanged and is rapidly degraded by the colonic microflora to produce HTyr, which can then be absorbed [
375]. OLE is preferentially degraded in vivo by lactic acid bacteria, such as
Lactobacillus and
Bifidobacterium species [
376], which use OLE as a carbon source and benefit from its metabolism. HTyr administration in high-fat-diet-fed mice was found to increase the concentration of the probiotic bacteria
Lactobacillus, specifically
L. johnsonii, and to not significantly change the ratio Firmicutes/Bacteroidetes (F/B), which was instead decreased in normal-diet-fed mice, indicating that HTyr may be beneficial for metabolism and may control F/B ratio [
377]. Dietary HTyr supplementation in high-fat-diet-fed mice also reduced the numbers of
Proteobacteria and
Deferribacteres, whose abundances have been associated with dysbiosis in hosts with metabolic or inflammatory disorders [
378], and promoted intestinal integrity reducing inflammation level [
377]. The ability of HTyr to stabilize the F/B ratio and restrain the increase of
Proteobacteria may also help controlling gut dysbiosis in inflammatory bowel disease patients, in which a decrease of Firmicutes and Bacteroidetes and an increase of
Proteobacteria are the most consistent hallmarks of disease [
379]. The beneficial effect of HTyr and OLE on gut microbiota is also boosted by their ability to inhibit pathogenic bacteria growth, either Gram-positive or Gram-negative, such as
E. coli [
72,
380].
The HTyr-induced modulation of colon microbiota was also correlated to its powerful antioxidant effect in oxidative-stressed mice [
178]. In fact, the HTyr-promoted decrease of the lipid peroxidation marker malondialdehyde (MDA) in the serum was correlated with the increase of the relative abundance of Firmicutes and
Lactobacillus and the decrease of Bacteroidetes, as an effect of HTyr administration [
178]. Moreover, HTyr decreased
Parabacteroides, whose abundance was associated with the progression of oxidative stress and inflammation [
178]. Interestingly, the HTyr-induced increase of Firmicutes positively correlated with a higher concentration of butyrate, one of the most important SCFAs, involved not only in maintaining intestinal function but also in reducing oxidative stress [
381]). Surprisingly, HTyr supplementation was demonstrated to be effective in relieving oxidative stress in mice also by increasing
Staphylococcales [
382], whose success as pathogens is due in part to their ability to mitigate endogenous and exogenous oxidative and nitrosative stress [
383]. The restrain of oxidative stress due to HTyr administration was also found to prevent fine dust (PM
2.
5)-induced adiposity and insulin resistance in adult mice [
384]. Once again, it was highlighted that gut microbiota may mediate the actions of HTyr, which was effective in increasing microbiota richness, reducing pathogenic bacteria, and reversing PM
2.
5-induced gut dysbiosis. In particular, the genus
Akkermansia, belonging to the Verrucomicrobia phylum and known to be beneficial for reducing weight gain and endotoxemia in mice and humans [
385], was exclusively abundant in HTyr-administered mice. In addition,
Ruminococcaceae and
Mycoplasmataceae, beneficial for glucose metabolism and abundant in long-living people, were enhanced by HTyr administration as well as
Prevotellaceae, which negatively correlated with lipid peroxidation biomarkers, thus contributing to the HTyr antioxidant effect [
384]. Besides HTyr, OLE was also found to ameliorate advanced-stage type 2 diabetes in mice by regulating gut microbiota [
386]. Consistent with the previously reported effects of HTyr administration, O LE was shown to significantly reduce the relative abundance of Bacteroidetes, including the
Parabacteroides genus, and increase that of Verrucomicrobia, including the
Akkermansia genus, confirming their importance in improving diabetes symptoms and insulin resistance [
386].
Altogether, the above studies highlighted that HTyr and OLE both act as prebiotics in gut microbiota modulation, favoring intestinal microbes’ homeostasis, even in the presence of chronic diseases. Their administration has been demonstrated to increase the diversity of gut microbiota, enhancing beneficial bacteria, and inhibiting pathogenic ones, thus impacting the health of multiple organs and systems. In fact, the well-established relationship between gut and CNS highlights the role of prebiotics in reducing the degree of oxidative stress and consequent progression of neurodegenerative and cardiovascular diseases and metabolic syndromes. For this reason, the maintenance of a condition of eubiosis is crucial to guarantee healthy aging, preventing and ameliorating several chronic pathologies. The potential therapeutic application of prebiotics supplied with the diet is increasingly investigated in order to keep optimal health during aging and also for the treatment of age-related disorders.