2. Molecular Mechanisms of Exosomes
2.1. Musculoskeletal Regeneration
With the aging of the global population, the incidence of musculoskeletal diseases is steadily increasing [
16]. Exosomes have gained great attention in musculoskeletal research due to their regenerative potential and minimally invasive nature (
Figure 1).
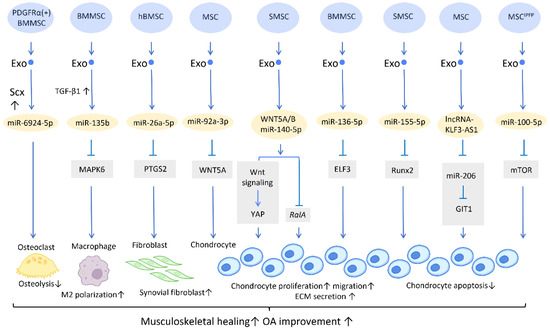
Figure 1. Molecular mechanisms of stem cell-derived exosomes in sport injuries and osteoarthritis. Stem cell-derived exosomes promote tissue healing, M2 polarization, fibroblast activation, chondrocyte proliferation and migration, and decreases osteolysis and chondrocyte apoptosis via transferring of diverse molecular components. An overall regenerative effect was demonstrated, which manifested as improved musculoskeletal healing and osteoarthritis recovery. Abbreviation: Scx: scleraxis; PTGS2: prostaglandin-endoperoxide synthase 2; YAP: Yes-associated protein; ELF3: E74-like factor 3; GIT1: G-protein-coupled receptor kinase interacting protein-1; hBMSC: human bone MSC; SMSC: synovial MSC; MSCIPFP: infrapatellar fat pad. ↑ represents upregulation; ↓ represents downregulation.
2.1.1. Sports Medicine and Musculoskeletal Rehabilitation
In contrast to the prevalent application of platelet-rich plasma (PRP) in clinical settings, the development of exosome-based therapies is still preliminary, and studies are largely preclinical. Recent research has gradually demonstrated the therapeutic efficacy of exosomes in musculoskeletal healing [
17,
18]. Exosomes derived from adipose-derived stem cells (ADSC-Exos), for example, have been shown to promote human rotator cuff healing by inhibiting muscle atrophy and degeneration, while also improving the histological properties of the torn tendon [
19]. This could be accomplished by increasing AMP-activated protein kinase (AMPK) signaling, and thus inhibiting Wnt/β-catenin activity [
20]. In addition, bone mesenchymal stem cells (BMMSCs)-derived exosomes (BMMSC-Exos) have been shown to stimulate the proliferation and differentiation of tendon stem/progenitor cells in vitro, increase the expression of mohawk, tenomodulin, and type I collagen, enhance the proliferation of local tendon stem/progenitor cells in vivo, and promote the mechanical properties of neotendons in the defect area of rat patellar tendons [
21].
Furthermore, a potential treatment for tendon–bone regeneration was proposed, utilizing exosomes derived from genetically modified BMMSCs. Scleraxis is a transcription factor that is believed to play an active role in tendon–bone repair. A study by Feng et al. demonstrated that local injection of miR-6924-5p-rich exosomes, derived from scleraxis-overexpressing PDGFRα (+) BMMSCs, leads to reduced osteolysis and improved healing strength [
22] (
Figure 1).
2.1.2. Osteoarthritis
The treatment of osteoarthritis (OA) has been challenging due to the low potential for spontaneous healing of cartilage tissue [
23].
In the application of exosomes in OA, several studies have proposed possible mechanisms behind its effect. It is well recognized that the proinflammatory cytokine IL-1β is one of the most influential players in cartilage destruction in OA [
24]. BMMSC-Exo injection significantly reduces IL-1β-mediated suppression of chondrocyte proliferation, and thereby inhibits the downregulation of anabolic markers COL2A1 and ACAN, as well as blocks the overexpression of catabolic markers MMP13 and ADAMTS5. This treatment reduced both cartilage damage and knee pain in rats with OA [
25]. In addition, exosomes derived from human embryonic stem cell-induced mesenchymal stem cells (ESC-MSCs) have shown therapeutic promise for the treatment of OA. In the presence of IL-1β, exosomes from ESC-MSCs were able to modulate chondrocytes by increasing the expression of the extracellular matrix protein collagen type II and decreasing the expression of the matrix degradation enzyme ADAMTS5. This effectively prevented cartilage destruction in the progression of OA [
26]. A study has shown that high expression of miR-135b in TGF-β1-stimulated BMMSC-Exos (BMMSC-Exos
TGF-β1) reduces pro-inflammatory factors and alleviates cartilage damage in OA. This is likely achieved by targeting MAPK6, which in turn boosts the M2 polarization of synovial macrophages [
27] (
Figure 1). Another study demonstrated that IL-1β-induced damage to synovial fibroblasts might be mitigated by exosomal miR-26a-5p, from human bone MSCs (hBMSCs), by hindering prostaglandin-endoperoxide synthase 2 (PTGS2) [
28] (
Figure 1).
WNT5A has a dual role, functioning in both chondrogenic differentiation and cartilage degradation, and it is involved in the destruction and degradation of cartilage in the formation of OA [
29]. Exosomal miR-92a-3p, released from MSCs, precluded cartilage destruction in an OA mouse model by downregulating WNT5A expression [
30] (
Figure 1). Furthermore, WNT5A and WNT5B were substantially expressed in synovial MSC exosomes (SMSC-Exos). WNT5A and WNT5B carried by exosomes stimulated Yes-associated protein (YAP) activation via an alternative Wnt signaling pathway, which contributed to the enhancement of chondrocyte proliferation and migration. Overexpression of miR-140-5p could block the side effect of YAP by suppressing
RalA, which rescued SOX9, aggrecan, and collagen type II expression and restored extracellular matrix (ECM) secretion [
31] (
Figure 1).
In the cartilage tissues of patients with traumatic OA, increased E74-like factor 3 (ELF3) expression and decreased miR-136-5p expression were found. Similar to the aforementioned miR-140-5p, exosomal miR-136-5p derived from BMMSCs was also shown to promote chondrocyte proliferation, migration, and ECM secretion by downregulating ELF3 expression, lessening MMP-1 levels, and intensifying collagen II, aggrecan, and SOX9 levels, thereby inhibiting cartilage degradation [
32] (
Figure 1). Furthermore, miR-155-5p-overexpressing SMSC-Exos might stimulate chondrocyte proliferation and migration while decreasing apoptosis, as well as increasing ECM secretion by targeting Runx2 to prevent OA [
33] (
Figure 1). Similarly, exosomal lncRNA-KLF3-AS1 generated from MSCs was reported to reduce chondrocyte apoptosis and stimulate chondrocyte proliferation, via sponging miR-206 to enhance G-protein-coupled receptor kinase interacting protein-1 (GIT1) expression, which attenuates IL-1β-induced chondrocyte injury [
34] (
Figure 1). Another study explored the effect of infrapatellar fat pad (IPFP) MSC-derived exosomes (MSC
IPFP-Exos) in OA. MiR-100-5p-abundant MSC
IPFP-Exos prevented damage to articular cartilage and lessened the severity of OA by hampering apoptosis of chondrocytes through the mTOR-autophagy signaling pathway [
35] (
Figure 1).
2.2. Wound Healing
Wound healing is a dynamic physiological process that may be roughly broken down into four stages: hemostasis, inflammation, proliferation, and remodeling. In this complicated process, numerous aspects, including angiogenesis, proliferation and differentiation of fibroblasts, and immunomodulation, play their roles in different stages of wound healing [
36]. The effect of exosomes on the healing of wounds has long been a subject of significant research and clinical interest. Studies show that extracellular signal-regulated kinase, STAT-3, and protein kinase B are just a few of the pathways that exosomes can target. These pathways are essential for aiding in and speeding up wound healing via upstream effectors, such as insulin-like growth factor-1, hepatocyte growth factor, nerve growth factor, and stromal cell-derived factor. Low quantities of TGF-β have been discovered in MSC-Exos in the umbilical cord, and, when exosomes are further laden with TGF-β cargo, they promote the remodeling of the matrix and vascularization [
37].
2.2.1. Stimulation of Angiogenesis
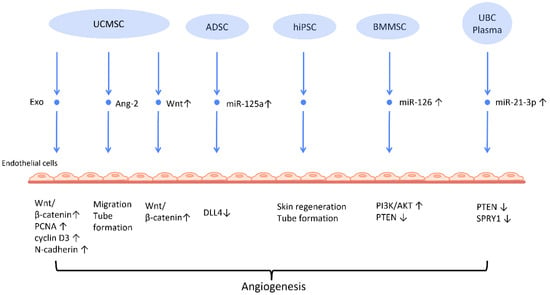
Neovascularization is a paramount process in wound healing and tissue regeneration (
Figure 2). HiPSC-MSC-Exos were first found to exhibit the ability to stimulate angiogenesis in human umbilical vein endothelial cells [
38] (
Figure 2). Further research indicates plausible mechanisms behind this finding. UCMSC-derived exosomes (UCMSC-Exos) transmitted angiopoietin-2 (Ang-2) to human umbilical vein endothelial cells [
39], thus enhancing their proliferative, migratory, and tube-forming capabilities. Additionally, another study reported that UCMSC-Exos also transmitted miR-21-3p to HUVEC, which increased angiogenesis and proliferation by inhibiting phosphatase and tensin homolog (PTEN), and sprouty homolog 1 (SPRY1) [
40] (
Figure 2). Exosomes derived from ADSCs-Exos can also trigger the proliferation and migration of vascular endothelial cells, thus promoting angiogenesis. In another study, it was found that transferring miR-125a from ADSC-Exos to endothelial cells increased angiogenesis by blocking the effect of delta-like 4 (DLL4), an inhibitor of angiogenesis [
41] (
Figure 2).
Figure 2. Molecular mechanisms of stem cell-derived exosomes in stimulation of angiogenesis. Stem cell-derived exosomes facilitate neovascularization by transmitting various angiogenic factors or miRNA to endothelial cells, leading to increased proliferation and migration of endothelial cells. Abbreviation: Exo: exosome; MSC: mesenchymal stem cell; UCMSC: human umbilical cord MSC; PCNA: proliferating cell nuclear antigen; ADSC: adipose-derived stem cells; DLL4: delta-like 4; hiPSC-MSC: human induced pluripotent stem cell-derived mesenchymal stem cells; BMSC: bone marrow-derived MSCs; Ang-2: angiopoietin-2; UBC: umbilical cord blood; PTEN: phosphatase and tensin homolog; SPRY1: sprouty homolog 1. ↑ represents upregulation; ↓ represents downregulation.
Another mechanism that has been often addressed in MSC-Exos is the Wnt/β-catenin signaling pathway, which plays a pivotal role in the proliferative phase of the wound healing process [
42]. The transfer of Wnt4 from UCMSC-Exos to endothelial cells activated the Wnt/β-catenin pathway and increased the number of angiogenesis-related factors, including proliferating cell nuclear antigen (PCNA), cyclin D3, and N-cadherin, to promote angiogenesis for the healing of second-degree burn injuries [
43]. In the rat skin burn model, UCMSC-Exos containing Wnt4 stimulated the Wnt/β-catenin pathway and the AKT pathway to increase angiogenesis and decrease heat stress-induced apoptosis, respectively [
44]. ADSC-Exos hindered the H
2O
2-induced apoptosis of HaCaT cells via stimulating the Wnt/β-catenin signaling pathway, which may be advantageous for cutaneous wound healing [
45].
The angiogenic efficacy of MSC-Exos has also been applied to diabetic wound healing, where hyperglycemia impairs angiogenesis and impedes the wound healing process [
46]. ADSC-Exos that overexpressed nuclear factor-E2-related factor 2 (Nrf2), a transcription factor that favors anti-oxidative properties, not only promoted proliferation of endothelial progenitor cells and angiopoiesis, but also increased granulation tissue, growth factor levels, and vascularization in diabetic mouse wounds while decreasing inflammatory and oxidative stress-related proteins [
47].
Recent investigations have found that exosomes derived from preconditioned MSC are more therapeutically efficacious than those obtained from MSC grown under standard conditions. By transferring miR-126, which downregulated PTEN and activated the PI3K/AKT signaling pathway [
48], BMMSC-Exos with deferoxamine, a hypoxia-inducing agent, strengthened the proliferative, migratory, and proangiogenic capacity of human umbilical vein endothelial cell and diabetic wound healing.
2.2.2. Proliferation and Collagen Synthesis of Fibroblast
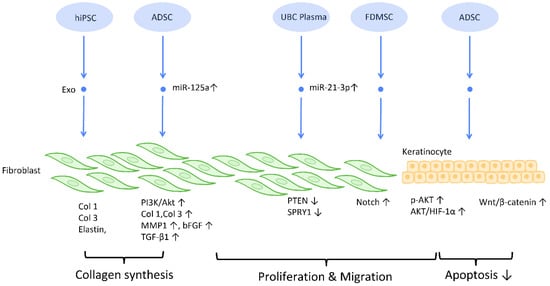
To establish an intact epidermal barrier, fibroblasts proliferate and produce ECM, which includes fibronectin, type 1 collagen, and type 3 collagen. Accumulating evidence suggests that MSC-Exos may be absorbed by fibroblasts and thereby stimulate collagen production (
Figure 3). HiPSC-MSC-Exos increased the production, migration, and secretion of type 1 collagen, type 3 collagen, and elastin by human dermal fibroblasts (DF) [
38]. ADSC-Exos transferred to DF enhanced collagen-related proteins, including type 1 collagen, type 3 collagen, MMP1, bFGF, and TGF-β1, and activated PI3K/Akt signaling to maximize collagen deposition [
49] (
Figure 3). In addition, exosomes produced from fetal dermal MSC-derived exosomes (FDMSC-Exos) augmented the secretion, migration, and proliferation of adult DF via activation of the Notch signaling pathway [
50] (
Figure 3).
Figure 3. Molecular mechanisms of stem cell-derived exosomes in regulation of fibroblast and keratinocyte. Stem cell-derived exosomes enhance the proliferation and collagen synthesis of fibroblasts, thereby improving the healing process. ADSCs are also shown to accelerate keratinocyte migration and proliferation, which contributes to augmented healing activity. Abbreviation: Exo: exosome; Col: collagen; MSC: mesenchymal stem cell; ADSC: adipose-derived stem cell; miR: micro-RNA; TGFβ: transforming growth factor-β; PI3K/Akt: phosphatidyl-inositol 3-kinase/serine-threonine kinase; MMP: matrix metalloproteinase. ↑ represents upregulation; ↓ represents downregulation.
2.2.3. Inhibition of Scar Formation
In the process of wound healing and tissue repair, the excessive accumulation of myofibroblasts and deposition of collagen results in aberrant scar formation. By controlling fibroblast transition and ECM remodeling, administrations of stem cell-generated exosomes have the potential to limit scar formation, and even accomplish scar eradication, striking a balance between therapeutic effects and aesthetic concerns (
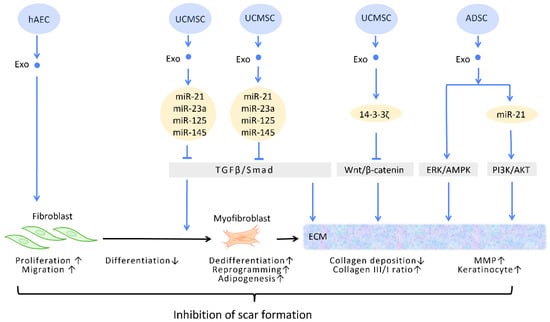
Figure 4) [
51].
Figure 4. Molecular mechanisms of stem cell-derived exosomes in inhibition of scar formation and promotion of ECM remodeling. In the process of tissue repair, excessive accumulation of myofibroblasts and deposition of collagen result in aberrant scar formation. By controlling fibroblast transition and ECM remodeling, administrations of stem cell-generated exosomes have the potential to limit scar formation. Abbreviation: ECM: extracellular matrix; ERK: extracellular signal-regulated kinase; MAPK: mitogen-activated protein kinase; PI3K/Akt: phosphatidyl-inositol 3-kinase/serine-threonine kinase; MMP: matrix metalloproteinase. ↑ represents upregulation; ↓ represents downregulation.
In the wound healing microenvironment, macrophages secrete TGF-β1, which initiates the differentiation of fibroblasts into myofibroblasts. Disrupting the TGF-β/Smad pathway to prevent TGF-β1-induced differentiation of fibroblasts into myofibroblasts is a straightforward strategy for minimizing fibrosis and scarring under these circumstances [
52] (
Figure 4). In this context, evidence suggested that UCMSC-Exos, enriched in miR-21, miR-23a, miR-125b, and miR-145, inhibited TGF-β/Smad2 and myofibroblast formation [
53] (
Figure 4).
2.2.4. Extracellular Matrix Remodeling
Accumulating evidence has suggested that exosomes are able to remodel and adjust the protein composition of the ECM, modifying the ECM into a more preferable condition throughout the process of wound healing [
54].
The ECM is composed of a variety of proteins, the most prevalent of which is collagen. Collagen possesses dual roles during different stages of wound healing. During the early phase of wound healing, stimulation of collagen synthesis is essential for restoration of wound strength, but excessive collagen during the late phase of wound healing results in scar formation [
55]. It has been reported that, aside from promoting the formation of myofibroblasts, the aforementioned TGF-β/Smad pathway is also involved in the regulation of collagen synthesis. The upregulation of the TGF-β/Smad pathway intensifies the expression of the
COLI2 gene during the early phase of wound healing but decreases collagen I deposition during the late phase [
53]. Through the targeting of this pathway, UCMSC-Exos promote favorable ECM composition throughout the healing process [
56] (
Figure 4).
Zhang et al. demonstrated that UCMSC-Exos transport 14-3-3ζ proteins and diminishes collagen deposition. 14-3-3ζ proteins facilitate the binding between YAP and phosphorylation-large tumor suppressor kinase, resulting in YAP phosphorylation and the blockage of Wnt/β-catenin signaling, which causes skin fibrosis during the remodeling phase of healing [
10] (
Figure 4).
Matrix metalloproteinases (MMPs) are components of the ECM that degrade excessive collagen. It has been suggested that ADSC-Exos trigger the ERK/MAPK pathway and augment the expression of MMP-3 [
57] (
Figure 4). Furthermore, Yang et al. revealed that miR-21 in ADSC-Exos regulates the PI3K/AKT pathway and elevates MMP-9 production, which accelerates HaCaT keratinocyte migration and proliferation [
51,
58] (
Figure 4).
2.3. Female Infertility
Impaired fertility is manifested as a declining pregnancy rate and poor pregnancy outcomes [
59]. By definition, infertility is the failure to establish a pregnancy within 12 months of regular sexual intercourse [
60]. Female reproductive disorders pose great threats to women’s health and eventually contribute to infertility [
61]. Stem cell-derived exosomes used in cell-free treatment have been shown to enhance the ovarian and uterine environments through their capacity for regeneration, which could potentially reverse female infertility.
2.3.1. Promoting Follicular Development
Several reproductive diseases are associated with disrupted follicle development. While ovarian aging is linked to a drop in the number and quality of oocytes, premature ovarian insufficiency, also known as premature ovarian failure, is a disease characterized by senescence of the ovaries in women under the age of 40 [
60]. The resulting follicular malfunction often manifests clinically as amenorrhea, estrogen deficiency, and hypergonadotropism [
62]. In addition, polycystic ovary syndrome (PCOS) is another common disease with ovulatory dysfunction among reproductive-aged females [
63].
The successful maturation of oocytes, which is pivotal for ovulation and pregnancy, is intimately connected with hormone regulation, granulosa function, and the ovarian environment [
64]. Since the number of oocytes defines the reserve of primordial follicles [
65], restoring the impaired follicular development is the key to improving ovulatory failure caused by ovarian reproductive disorders. Premature ovarian insufficiency is the most frequently adopted model for this line of research. In the vast majority of studies, anti-apoptosis and adjustment of sex hormone levels are hypothesized to be the underlying mechanisms of exosome therapy.
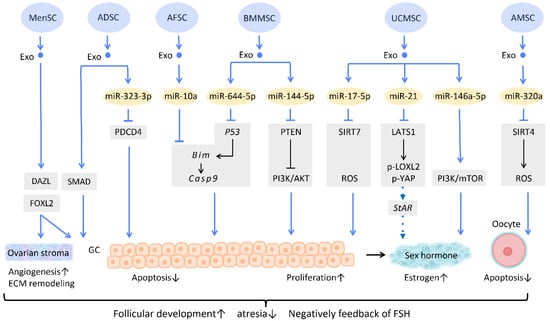
Several researchers have delved into the molecular pathway of how stem cell-derived exosomes exert their ability to inhibit apoptosis through their cargo (
Figure 5). Human ADSC-Exos were found to repress apoptosis of human granulosa cells by regulating the SMAD pathway, which elevates mRNA and protein expression of SMAD2, SMAD3, and SMAD5 to a normal level [
66] (
Figure 5). Furthermore, exosomes derived from ADSC transfected with miR-323-3p eliminate the pro-apoptotic action of programmed cell death protein 4 (PDCD4) in PCOS cumulus cells [
63] (
Figure 5). Exosomes derived from amniotic fluid stem cells (AFSC-Exo) deliver miR-10a and reduce follicular atresia by targeting
Bim and downregulating the apoptotic factor
Casp9 [
67] (
Figure 5). In parallel, exosomes collected from BMMSC-Exos, carrying miR-644-5p, impede apoptosis of ovarian GCs through targeting
p53 [
68] (
Figure 5), the upstream of
Bim. Meanwhile, BMMSC-Exos are capable of activating the PI3K/AKT pathway in granulosa cells by suppressing PTEN, an inhibitor of this signaling pathway [
69] (
Figure 5). On the other hand, UCMSC-Exos highly express miR-17-5p, inhibiting sirtuin 7 (SIRT7) and its downstream genes
PARP1,
γH2AX, and
XRCC6. This results in a decreased accumulation of reactive oxygen species (ROS) in granulosa cells, which has an anti-apoptotic effect [
70] (
Figure 5).
Figure 5. Molecular mechanisms of stem cell-derived exosomes in follicular development. Follicular malfunction is the critical reason of ovarian reproductive disorders. With the help of stem cell-derived exosomes, follicular growth and sex hormone levels could be restored, and ovarian microenvironment could be rejuvenated. Abbreviation: Exo: exosome; SC: stem cell/stromal cell; MSC: mesenchymal stem cell; MenSC: menstrual blood-derived SC; ADSC: adipose-derived stem cell; AFSC: amniotic fluid stem cell-derived; BMMSC: bone marrow-derived MSC; UCMSC: umbilical cord MSC; AMSC: amniotic MSC; miR: micro-RNA; DAZL: deleted in azoospermia like; FOXL2: forkhead box L2; PDCD4: programmed cell death protein 4; PI3K/Akt: phosphatidyl-inositol 3-kinase/serine-threonine kinase; SIRT: sirtuin; ROS: reactive oxygen species; LATS1: large tumor suppressor 1; LOXL2: lysyl oxidase-like 2; YAP: Yes-associated protein; StAR: steroidogenic acute regulatory gene; GC: granulosa cell; ECM: extracellular matrix. ↑ represents upregulation; ↓ represents downregulation.
Regarding regulation of sex hormone levels, one study revealed that exosomal miR-21 is involved in enhancing estrogen secretion in hUCMSCs (
Figure 5). Further effort was made to clarify the molecular mechanism underlying this observation. Exosomal miR-21 was found to downregulate large tumor suppressor 1 (LATS1), which belongs to the Hippo pathway, thereby mitigating phosphorylation of lysyl oxidase-like 2 (LOXL2) and YAP. In this way, YAP protein could bind to the promotor of the steroidogenic acute regulatory (
StAR) gene, thereby elevating
StAR expression and promoting estrogen secretion in ovarian granulosa cells [
71] (
Figure 5).
Aside from acting on granulosa cells, exosomes also have an effect on oocytes (
Figure 5). For instance, the mechanism of miR-320a, from human amniotic MSC-derived exosome (AMSC-Exo), inhibits oocyte apoptosis by suppressing SIRT4 signaling and reducing ROS levels [
72] (
Figure 5), an effect which is similar to that of miR-17-5p from BMMSC-Exos. In the model of age-related fertile retardation, UCMSC-Exos carries miR-146a-5p, which triggers the PI3K/mTOR pathway, improving oocyte quality and accelerating follicular development [
65] (
Figure 5). By means of promoting follicular development and obliterating atresia, stem cell-derived exosomes restore ovarian function and negatively feedback on follicle-stimulating hormone (FSH) levels.
In addition to the above-mentioned mechanisms, several studies indicate that exosome therapy can adjust ovarian stroma into a more favorable microenvironment. Zhang et al. showed that exosomes from menstrual blood-derived stromal cells not only stimulated the expression of early follicle markers Deleted in Azoospermia-like (DAZL) and Forkhead Box L2 (FOXL2), but also regulated the composition of the ECM. Enhanced expression of collagen IV, FN1, and laminin in the ovarian stroma was noted; the latter two were also observed in granulosa cells, and are involved in follicular growth [
64] (
Figure 5).
2.3.2. Regeneration of Damaged Endometrium
Being the implant sites of blastocytes, endometrium undoubtedly plays a crucial role in female pregnancy. Asherman’s syndrome, also called intrauterine adhesion (IUA), is characterized by poor glandular epithelial growth and vascular development from scarring, and fibrosis primarily caused by artificial trauma, repeated endometritis, and infection [
59,
73,
74]. Endometrial fibrosis causes a loss of regeneration competence in functional endometrium, which eventually leads to secondary infertility, the most common type of infertility worldwide [
60,
75].
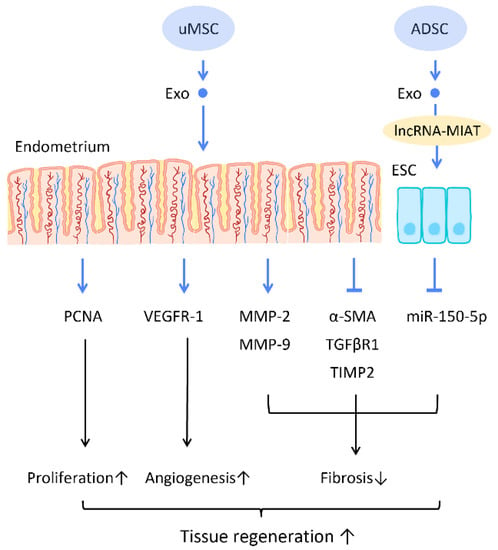
Recent research has found that the administration of stem cell-derived exosomes has similar effects to PRP in endometrial fibrosis and AS models, including promoting cellular proliferation, differentiation, functional recovery, angiogenesis, anti-inflammation, and anti-fibrosis. Saribas et al. utilized uterus-derived MSCs-exosomes (uMSC-Exo) in AS models and found increased proliferation, vascularization, and reduced fibrosis in uterine tissue. The effect was confirmed by significantly elevated proliferating cell nuclear antigen (PCNA), VEGFR-1, MMP-2, MMP-9, and declined TIMP-2 [
76] (
Figure 6). Concerning specific molecular interactions between exosomes and endometrial cells, Shao et al. established a negative regulatory relationship between lncRNAs and miRNAs. They elucidated that ADSC-Exos transfer lncRNA-MIAT by targeting miR-150-5p in endometrial epithelial cells, and alleviate fibrosis of endometrial tissue with a higher expression of cytokeratin 19 and a lower expression of fibrosis markers α-SMA and TGFβR1 [
75] (
Figure 6).
Figure 6. Molecular mechanisms of stem cell-derived exosomes regeneration of damaged endometrium. The administration of stem cell-derived exosomes is beneficial in female pregnancy, which alleviates endometrial fibrosis and enhances the regeneration competence in endometrial tissues. Abbreviation: Exo: exosome; MSC: mesenchymal stem cell; uMSC: uterus-derived SC; ADSC: adipose-derived stem cell; lncRNA: long noncoding chain RNA; VEGF: vascular endothelial growth factor; VEGFR: VEGF receptor; MMP: matrix metalloproteinase; TIMP: tissue inhibitor of metalloproteinase; α-SMA: alpha smooth muscle actin; miR: micro-RNA. ↑ represents upregulation; ↓ represents downregulation.