Carboxyl/cholinesterases (CCEs) represent a family of enzymes distributed in many organisms, including insects. Despite their relatively simple catalyzed hydrolysis reaction, CCEs facilitate insects’ adaptation to chemical signals and stressors from the environment through various trajectories, including developing pesticide resistance, facilitating the adaptation of insects to their host plants, and manipulating insect behaviors. The CCE-mediated mechanisms of pesticide resistance to organophosphate, carbamate, or pyrethroid pesticides comprise enhanced metabolism, the sequestration of pesticides to prevent them from reaching their target sites, or conformational changes in target sites to prevent pesticides from binding. In addition, CCEs aid in the adaptation to chemical signals through the olfactory system by degrading insect semiochemicals.
1. Introduction
Insects have evolved sophisticated mechanisms to adapt to various xenobiotics, such as plant allelochemicals, insect odors and pheromones, pesticides, and industrial pollutants [
1,
2,
3]. Metabolic detoxification is a multi-phase process involving the enzymatic degradation and conjugation of lipophilic compounds to water-soluble, excretable metabolites [
3,
4,
5,
6]. In Phase I, cytochrome P450 monooxygenases (P450s) and carboxyl/cholinesterases (CCEs) convert xenobiotics from more lipophilic compounds to more hydrophilic products. In Phase II, glutathione S-transferases (GSTs) and/or UDP-glycotransferases (UGTs) further conjugate the xenobiotic metabolites or Phase I products and prepare them for excretion. In Phase III, the products can be excreted through cellular transporters, such as ATP-binding cassette (ABC) transporters [
5,
7,
8].
CCEs constitute a multigene family of α/β-hydrolase fold enzymes that are distributed in numerous organisms including insects, mammals, plants, and microorganisms [
9,
10,
11,
12,
13]. As Phase I enzymes, insect CCEs hydrolyze structurally diverse xenobiotics containing ester or amide bonds that consist of pesticides, insect and plant odors, insect pheromones and hormones, as well as environmental toxicants [
9,
14,
15,
16]. Due to their versatile capabilities in hydrolytic biotransformation, insect CCEs play vital roles in the development of pesticide resistance, facilitating the adaptation of insects to their host plants and manipulating insect behaviors through olfaction [
9,
17,
18].
2. CCE Classification and Structural Characteristics
2.1. Classification of Insect CCEs
Insect CCEs are divided into 14 clades in three classes based on their physiological and biochemical functions: dietary/xenobiotic detoxification (clades A–C), pheromone/hormone processing (clades D–H), and neuro/developmental functions (clades I–N) [
9,
19,
20]. The dietary detoxification class of CCEs is of the highest quantity and sequence variation among species. CCEs in this class are identified as intracellular catalytically active enzymes and belong to α-esterases [
19,
20]. There are three major clades in this class, the A–C clades. Most insect species have the clade B CCEs (microsomal α-esterases) in their genomes [
20].
There are five major clades in the secreted, catalytically active pheromone/hormone processing class: integument esterases (Clade D), β-esterases (Clade E), juvenile hormone esterases (JHEs) (Clades F and G), as well as glutactin and similar enzymes (Clade H) [
19,
20]. Clade D is composed of one to several integument esterases or semiochemical esterases in each insect genome. The functions of integument esterases are implicated in pheromone and other semiochemically triggered signaling processes [
15,
19,
21,
22,
23,
24]. Recent studies have suggested that some insect integument esterases may contribute to xenobiotic detoxification [
14,
25]. β-esterases in Clade E exhibit a wide range of functions that have been extensively studied [
19,
20].
The neuro/developmental class include the acetylcholinesterase (AChE) and five noncatalytic clades (Clades I-N) [
20,
40]. Aside from AChEs, many of the neuro/developmental class are membrane-bound and fall under the subclassification CLAMs (cholinesterase-like adhesion molecules), of which one or more residues of the catalytic triad are absent [
41]. CLAMs include neuroligins, glioactins, neurotactins and glutactins, with the latter two only found in insects [
41,
42]. The major function of AChEs in insects is the hydrolysis of the neurotransmitter, acetylcholine (ACh), after it binds to the Ach receptor (AChR) at the cholinergic synapses [
19]. Mutations on AChE can inhibit OP and carbamate toxicities and have been linked with OP and carbamate resistance in many insect species [
43,
44,
45,
46,
47]. Except for AChEs, there are few detailed studies on other genes in the neuro/developmental class in insects, mainly in
D. melanogaster [
19,
42,
48,
49].
2.2. Structural Characteristics of Insect CCEs
All currently characterized insect CCEs contain the canonical α/β hydrolase fold backbone structure, with six interspersed α-folds distributed about a core β-sheet that contains eight β-strands, with seven of the core β-strands running in the parallel direction and one β-strand in anti-parallel formation [
50]. Additional β-strands, helices, and coils surround the core α/β hydrolase fold but vary across clades. This conserved backbone maintains the position of the enzyme’s active site: the catalytic triad and the oxyanion hole. In turn, the catalytic triad is the essential component in the CCE hydrolysis reaction [
9,
13]. The six insect CCE crystal structures resolved to date have a catalytic triad consisting of a serine, a glutamate and a histidine, except for Est6 from
D. melanogaster with an aspartate in place of the glutamate (
Table 2) [
37,
51,
52,
53,
54]. During the catalyzed ester hydrolysis reaction, the ester bond of the substrate is cleaved, resulting in the formation of an alcohol and a carboxylic acid product. To start the reaction, the acidic residue of the catalytic triad (Glu or Asp) is hydrogen bonded to the His, stabilizing it as a general base and allowing it to deprotonate the nucleophilic Serine. The nucleophilic Serine then attacks the carbonyl carbon of the bound ester substrate, forming a tetrahedral intermediate that is, in turn, stabilized by the oxyanion hole. Next, the tetrahedral intermediate collapses, deprotonating the Histidine and releasing the alcohol product, and forming the acyl-enzyme intermediate. The Histidine then deprotonates a water molecule, activating it to attack the carbonyl carbon of the acyl-enzyme intermediate, releasing the acid product and returning the enzyme to its resting state. Often, the structure of the CCE active site allows for the metabolism of many substrates, but preferences in CCEs for specific xenobiotics are observed [
9,
13,
37].
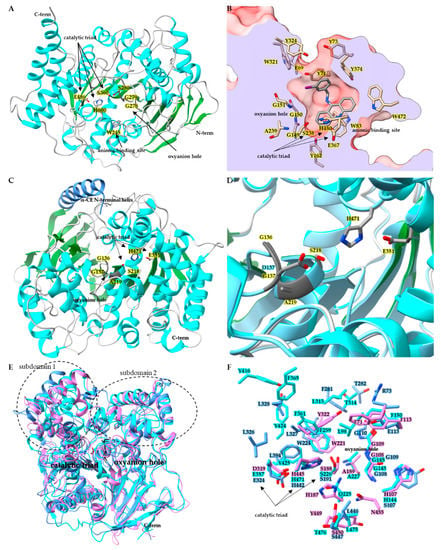
Belonging to the neuro/developmental class, insect AChE (E.C. 3.1.1.7) contains the core α/β hydrolase that is conserved across species (
Figure 1A) [
55]. A comparison of superimposed AChE catalytic domains from
Anopheles gambiae (
AgAChE PDB:6ARX),
D. melanogaster (
DmAChE:PDB 1QO9),
Homo sapiens (
hAChE:PDB 4EY4),
Mus musculus (
mAChE PDB:2HA2) and
Torpedo californica (
TcAChE PDB:2WG2) revealed α-carbon backbone root-mean-square-deviation (RMSD) ranges from 1.1 Å to 1.9 Å [
55,
56]. A key structural characteristic of AChE is the active site gorge, a channel starting at the surface of the AChE and extending deep into the protein and terminating at the catalytic/acetylcholine binding site (
Figure 1B) [
52]. In the
DmAChE:PDB 1QON crystal structure with inhibitor (1,2,3,4-tetrahydro-N-(3-iodophenyl-methyl)-9-acridinamine) bound in the acetylcholine binding site, there is a 17.7 Å distance from the hydroxyl of Tyr73 at the entrance of the active site gorge to the hydroxyl of Tyr162 at the bottom of the gorge, below the catalytic triad. For
AgAChE PDB:6ARY, the distance from the γ
2 carbon of Val235 (positionally equivalent to Tyr73 in
DmAChE) at the gorge entrance to the hydroxyl of Tyr291 (equivalent to Tyr162 in
DmAChE) is 17.9 Å [
52,
56]. Much of the AChE active site gorge is lined with aromatic residues thought to aid the delivery of the substrate to the catalytic site containing the catalytic triad, oxyanion hole and the conserved anionic binding site tryptophan [
56]. In
DmAChE, the catalytic triad is made up of Ser238, His480, and Glu367; adjacent to the triad is the oxyanion hole composed of Gly150, Gly151, and Ala239 backbone amides, and the conserved anionic binding site Trp83 (
Figure 1B) [
52]. Multiple instances of a G280S mutation in
AgAChE located in the oxyanion hole have been documented (
Figure 1A) [
56]. Structural analysis by Cheung et al. revealed that the G280S mutation in the oxyanion hole results in steric crowding of the acetylcholine binding site, with the hydroxyl of S280 pointing towards the catalytic triad, and as a result steric crowding leads to the loss of target site sensitivity towards large OP and carbamate insecticides [
56].
Figure 1. Structures of insect CCEs. (A) Ribbon diagram of acetylcholinesterase (PDB: 6ARX) from Anopheles gambiae. Side chains for key active site residues are shown and colored by element: catalytic triad Ser360, Glu486, His600, and oxyanion hole residues Gly278, Gly279, Gly280Ser mutation, and the anionic binding site Trp245. (B) Surface representation of acetylcholine binding pocket of DmAChE PDB:1QON, clipped to show active site gorge. Active site gorge residues are colored by heteroatom and the bound inhibitor 9-(3-ioodobenzylamino)-1,2,3,4-tetrahdroacridine is colored by element. (C) Ribbon diagram of wild-type LcαE7 PDB:4FNG, showing catalytic triad side chains colored by element, and oxyanion hole colored dark gray. (D) Active site of LcαE7, wild-type structure PDB:4FNG is superimposed with mutant Gly137Asp PDB:5C8V. (E) Superimposed ribbon diagrams of juvenile hormone esterase MsJHE PDB:2FJ0 (colored in cyan), with β-esterases CqESTβ2 PDB:5W1U (colored in cornflower blue) and DmEST6 PDB:5THM (colored in orchid). Catalytic triad residues sidechains are shown and the oxyanion hole portion of the ribbon diagram is colored black. (F) Zoomed in binding pocket residues for the superimposed MsJHE PDB:2FJ0, CqESTβ2 PDB:5W1U and DmEST6 PDB:5THM structures with residues colored by heteroatom.
Insect CCEs are known to play multiple physiological roles, such as hormone, neurotransmitter, pheromone, odorant, lipid ester, and insecticide degradation [
51,
73,
74,
75]. Multiple instances of insecticide resistance have been reported resulting from overexpression and/or mutation events in α-esterases [
51,
74]. The inactivation of esterase enzymes can occur upon exposure to OPs via phosphorylation of the active site serine [
76]. Mutations that alter the active site can lead to the gained ability of the enzyme to overcome inactivation upon exposure to OPs. A notable mutation has been found to occur in an α-esterase belonging to the dietary/xenobiotic detoxification class (
Figure 1C,D) [
74]. The G137D mutation found in sheep blowfly
Lucilia cuprina α-esterase
LcαE7 caused gaining of function hydrolysis activity toward the OP chlorfenvinphos, and in turn resulted in lost activity towards model substrates α-naphthyl acetate, p-nitrophenyl acetate, and methylthiobutyrate (
Figure 1D) [
74].
The hormone/pheromone processing class contains the insect β-esterase and juvenile hormone esterase [
37,
53,
73]. Crystal structures for
CqESTβ2 (PDB:5W1U),
DmEST6 (PDB:5THM), and
MsJHE (PDB:2FJ0) revealed that the hormone/pheromone processing class exhibits the typical esterase structural elements (
Figure 1E and 1F). When the structures for
CqESTβ2 (PDB:5W1U),
DmEST6 (PDB:5THM) and
MsJHE (PDB:2FJ0) are superposed with
DmAChE PDB:1QON, the largest α-carbon RMSDs are observed in N and C terminal regions, along with the subdomain 1 and subdomain 2 regions adjacent to the catalytic triad that play a dominant role in defining the size and shape of the binding pocket and thus the substrate specificity of CCEs [
37,
52,
53,
73].
3. Dynamic Rules of Insect CCEs in Chemical Adaptation
3.1. CCE-Mediated Insecticide Resistance
Insect CCE, cytochrome P450 and GST enzyme families are commonly implicated in the development of insecticide resistance [
82]. Most enhanced CCE mediated detoxification is detected in OP and/or carbamate resistant populations. In some cases, enhanced CCE detoxification is also involved in the resistance to pyrethroids, by hydrolyzing or sequestering insecticides before they reach their target: voltage-gated sodium channels [
83]. The mechanisms of CCE-mediated resistance to these insecticides include quantitative changes (enhanced CCE gene expression and enhanced CCE activities) and qualitative changes (mutations occur in the active sites) [
17,
84,
85]. OPs and carbamates are degraded through a hydrolysis reaction similar to CCE natural substrate hydrolysis. Qualitative changes in CCEs can enhance OP metabolism through a mutation that increases OP hydrolysis ability, and in turn reduces a CCE’s natural substrate hydrolysis activity. Quantitative changes arise through an enhanced expression of a CCE that can bind an insecticide, but with little or no hydrolysis activity (i.e., sequestration) [
53,
85]. The enhanced CCE expression can occur through mutations in the regulatory sequence (
cis- or
trans-), or gene amplification. The enhanced CCE expression (quantitative changes) can also be combined with mutations in the coding sequence (qualitative changes) that cause boosted metabolism of insecticides or increased binding affinity, resulting in higher levels of insecticide resistance [
17]. Insensitivity of AChE is shown to arise through one or more mutations in the
Ace genes, which encode insect AChEs. Multiple mutations often lead to greater insensitivity and resistance than one mutation [
86,
87].
Peach-potato aphid (
Myzus persicae) variants showed cross resistance to OPs, carbamates and pyrethroids by upregulating the expression of β-esterases E4 or FE4 (different variants boost copies of E4 or FE4). Resistant variants had as many as 80 more copies of the gene at the genomic DNA level than the susceptible variant, with no change in the enzyme coding sequence [
88,
89]. E4 could hydrolyze dimethyl and diethyl OPs at a slow rate, resulting in hydrolysis of a small fraction of OPs. While the major role of E4 involved in OP resistance was through sequestration. E4 hydrolyzed carbamates at even slower rates, which was consistent with the relative resistance level that
M. persicae had for both insecticide classes. Interestingly, E4 was also able to hydrolyze the
trans-isomers of the pyrethroid insecticide permethrin, but not
cis-isomers, and at a significantly lower rate than OPs and carbamates [
89].
The fungus gnat
Bradysia odoriphaga, which in its larval form is a crop pest of the Chinese chive, has been known to develop insecticide resistance in field populations. The CCE
BoαE1 of the dietary/detoxification class, isolated from a laboratory colony, was transcriptionally upregulated after malathion exposure and showed hydrolysis capability for malathion. The induced upregulation of
BoαE1 was suggested to contribute to the resistance found in field populations [
96]. In
C. quinquefasciatus, a CCE
Cqestβ2 was upregulated and played a major role in OP resistance through acting as a “sponge” to intercept insecticides before reaching their target. The stopped-flow kinetic analysis showed that a covalent intermediate formed after the rapid OP binding of
Cqestβ2 with high affinity, resulting in irreversible and prolonged enzyme inhibition [
53]. By contrast, a duplication and series of mutations in a malathion resistant
Culex tarsalis generated a new malathion carboxylesterase (MCE I) that could hydrolyze malathion 18 times faster than MCE II. MCE II was in both the susceptible and resistant strains and neither MCE I nor MCE II showed higher expression, indicating that the insecticide resistance was due to qualitative changes but not quantitative changes [
97].
3.2. Metabolism of Plant Allelochemicals
CCEs combined with GSTs and Cytochrome P450s are three major superfamilies of detoxification genes involved in plant allelochemical detoxification which facilitate insect host plant adaptation [
2,
107,
108,
109,
110,
111,
112]. Cytochrome P450s are well known to play important roles in plant allelochemical metabolism and have been broadly studied [
17,
113,
114]. However, the involvement of specific CCE genes in the metabolism of plant allelochemicals is rare and requires further investigation. Most studies focus on a comparison of total esterase levels in response to different plant diets, and in some cases how this affects susceptibility to insecticides.
The western tiger swallowtail
Papilio rutulus larvae were tolerant of diets containing phenolic glycosides tremulacin and salicortin, both of which were found in one of their host plants, quaking aspen [
115]. When fed phenolic glycosides in conjunction with an esterase inhibitor, S,S,S-tributylphosphorotrithioate (DEF), the growth and survival of larvae were reduced by half compared to larvae fed phenolic glycosides alone and significantly reduced compared to larvae fed DEF alone, suggesting the involvement of CCEs in the metabolism of phenolic glycosides [
115]. In another species, Spongy moth (
Lymantria dispar), feeding on a diet with added phenolic glycosides (2 or 4% wet weight) induced both CCE and GST activity in the midgut of the larvae, indicating CCEs facilitating the host plant adaptation of Spongy moth [
116,
117]. In
Sitobion avenae, CCE activity was induced by indole alkaloid gramine. The increase in CCE activity was positively correlated with the concentration of gramine, suggesting that CCE may contribute to the host allelochemical adaptation [
118].
3.3. Odorant Degradation in the Olfactory System
Olfaction plays vital roles in insect communication. One of the major olfactory organs in insects is the antenna, which detects volatile semiochemicals (e.g., plant volatiles, pheromones, insect odors) from the external environment for host plant location, mating, and avoiding predators [
15,
122]. After volatile semiochemicals bind to odorant receptors, triggering signal transduction, insect olfactory pathways have evolved to include a dynamic process of signal inactivation by odorant-degrading enzymes (ODEs) [
18]. Highly effective ODEs are required for degrading odorants and pheromones rapidly [
123]. So far, there are many ODEs identified in the insect olfactory system, including cytochrome P450s, GSTs, CCEs, UGTs, aldehyde oxidases, epoxide hydrolases, and alcohol dehydrogenase [
15,
124,
125,
126]. The first odorant-degrading enzyme (ODE) discovered in insects was a CCE located in the pheromone sensitive sensilla in the antenna of a male silk moth,
A. polyphemus, in 1981 [
21]. In addition to the first ODE, two more CCEs were also found to metabolize pheromones; one located in the wing scales [
24], and the other located in integument tissues of both
A. polyphemus sexes [
21].
CCEs represent the first ODEs discovered to degrade insect pheromones and plant odors and remain the most promising ODE candidates. CCEs that are involved in odorant degrading mainly belong to β-esterases, integument esterases, or mitochondrial and cytosolic CCEs. In
Spodoptera littoralis (Egyptian cotton leafworm), SICXE10 and SICXE7, two CCEs predominantly expressed in olfactory sensilla, both hydrolyze the female-produced sex pheromones (Z9E11-14:Ac and Z9E12-14:Ac) and the green leaf volatile ((Z)-3-hexenyl acetate) in vitro. SICXE10 and SICXE7 were shown to be expressed in pheromone-sensitive sensilla and short sensilla, and both esterases hydrolyzed Z9E11-14:Ac, Z9E12-14:Ac, and (Z)-3-hexenyl acetate, suggesting that these two CCEs play a role in degrading pheromone and plant odorants [
127,
128].
This entry is adapted from the peer-reviewed paper 10.3390/insects14020194