Unlike in animals, dioecy among flowering plants is a rare phenomenon. The vast majority of angiosperm species have a bisexual flower that combines male (androecium) and female (gynoecium) reproductive organs. However, about a quarter of species have dioecious flowers, which can be located within the same plant (monoecious) or on different plants (dioecious). In recent years, the development of sequencing techniques, bioinformatics, and molecular biology has led to an increase in interest in the sex determination mechanisms among plants. It is noteworthy that a significant number of dioecious plants have economic value. At the same time, dioeciousness often complicates the growing process. This fact increases the relevance of studies on dioecious crops. In this review, we attempt to summarize the current information on sex chromosomes and the mechanisms of sex determination in dioecious plants, concentrating on species with agricultural importance.
- dioeciousness
- flowering plants
- monoecious plant
- dioecious plant
- sex chromosomes
1. Introduction
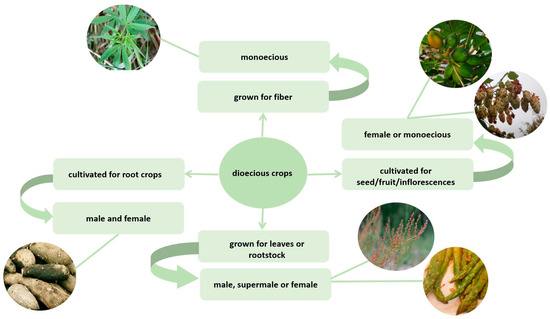
2. Features of the Dioecious Crop Cultivation
2.1. Sex Chromosomes and Sex Determination in Herbaceous Crops
2.1.1. Asparagus
2.1.2. Sorrel
The Rumex genus is one of the model groups for studying sex determination in plants [72]. Therefore, sex determination and sex chromosomes are widely studied in a number of Rumex species. Among these species, there are various systems for determining sex and sex chromosomes (completely hermaphroditic species, monoecious species with XY chromosomes and an active Y chromosome, and species with X:A balance sex determination, including multichromosomal species with the XY1Y2 system). Species such as R. acetosa, R. acetosella, R. hastatalus, and R. suffruticosus are dioecious and widely studied. The R. acetosa karyotype consists of 12 autosomes and sex chromosomes (XX in female plants and the XY1Y2 multichromosomal system in male plants) [73,74,75,76]. The sex chromosomes are the largest in the karyotype. Two Y chromosomes contain 26% of the total DNA. Sorrel Y chromosomes are heterochromatic and rich in repetitive DNA sequences [77]. In 1999, a group of tandem repeats specific to male sex chromosomes were isolated and called RAYSI (Rumex acetosa Y-chromosome-specific I) [78]. After that, a number of satellite DNA repeats specific for sorrel Y chromosomes were described. The repeat sequences of RAYSII and RAYSIII are AT-rich and show about 60% homology with RAYSI [73]. Repeat satDNA RAE180 is localized on two Y chromosomes and is also present on a pair of autosomes. [79].
2.1.3. Spinacia
2.1.4. Hemp
2.1.5. Betel
Betel rarely blooms, but there have been reports of economically significant differences in the varieties of male and female plants [30]. There is very little information about the chromosome numbers and the level of ploidy of this species. The main number of chromosomes of this species is x = 13; however, polyploid rows are reported in this culture. Apparently, plants with a high level of ploidy are mainly grown. It was shown that female plants are tetraploid and have 52 (2n = 4x = 52) chromosomes in their karyotype, while male plants are triploid and have 39 chromosomes (2n = 3x = 39) [119]. Sex markers have been created for betel plants, mainly to accompany the breeding process, which is currently not very developed due to the complexity in the biology of the culture [30,120,121].
2.1.6. Yams
Yams (Dioscorea rotundata) are one of the most important crops in a number of countries, primarily in the African continent, where Nigeria, Ghana, Côte d’Ivoire, and Benin are considered leaders in the production of yams [46]. The plant is grown for the production of tubers, which are then used for food. Normally, D. rotundata is a dioecious species; however, cases of monoecious occurrence have been observed. Despite the huge role of this crop in African countries, the selection of this species is difficult, largely due to its dioecious nature and problems with flowering (it is predominantly a vegetatively propagated crop) [122,123]. In addition, a change in the sex of plants under the influence of the environment is repeatedly observed during the growing season, which also complicates the breeding process of this crop [124]. The identification of sex chromosomes in yams is also difficult [125]. However, using modern sequencing techniques, the ZZ/ZW sex determination system has been demonstrated [125]. In 2020, the first yam genome obtained as long reads using the Oxford Nanopore Technologies technique was presented [126].
2.2. Sex Chromosomes and Sex Determination of Perennial Tree Crops Grown for Fruits
2.2.1. Papaya
The papaya sex chromosomes were the second to be fully sequenced after the human sex chromosomes [137]. They are evolutionarily young and outwardly practically homomorphic [11,138]. One of the features of papaya is the presence of hermaphroditic plants—in fact, it is a trioecious species, not a dioecious species. There are two types of Y chromosomes in this species. The first type is represented by the Y chromosome, which is found in male plants. The second type is represented by the Yh chromosome, which causes the hermaphroditism of flowers. At the same time, the YY, YYh, and YhYh genotypes are lethal [33,139]. The non-recombining region on the Y chromosome of papaya was formed as a result of two inversions of retrotransposons and is 8.1 Mb, while the size of the analogous region of the X chromosome is 3.5 Mb. All genes in non-recombining regions have been annotated [140]. Wild papaya is generally dioecious but cultivated as a rule of the gynodioecy cultivar [131]. It has been shown that evolutionarily hermaphroditic plants have been fixed for about 4000 years during domestication [141]. In addition to differences in the ease of cultivation (when growing hermaphroditic plants, male pollinating plants that do not produce fruits are not needed) between female and hermaphroditic plants, there is dimorphism in the shape of the fruits. Fruits from hermaphrodite flowers are more elongated and commercial producers prefer them, rejecting the more rounded fruits of female plants [137]. However, plants can change the sex of their flowers during the growing process. Hermaphroditic flowers can stop the development of carpels, turning into functionally male flowers under the influence of environmental factors such as drought or high temperatures [142,143,144].
2.2.2. Vitis
It has been shown that the emergence of hermaphroditic forms occurred about 6000–8000 years ago at the time of the domestication of wild grapes [5,148,149]. Like papaya, grapes have three sex-determining loci—Y (M) is male, X (F) is female, and Yh (H) is hermaphroditic. Unlike papaya, the HH, HM, or MM genotypes are viable in grapes. Three sex-linked grape genes (ViviPLATZ, VviFSEX, and APRT3) have been described, and the expression of each differs in hermaphroditic plants [149].
2.2.3. Diospyros
It is an autopolyploid species with 2n = 90 or 135 chromosomes (2n = 6x; 2n = 9x; x = 15) [152]. Despite the presence of at least one Y chromosome in the karyotype, this species is usually monoecious or has completely female plants, with rare male flowers [34]. Recent studies have shown that in the dioecious diploid species D. lotius L. (a close relative of D. kaki), the OGI gene located on the Y chromosome is responsible for sex formation. It encodes a small RNA that targets the autosomal MeGl gene, regulating anther fertility in a dose-dependent manner. In polyploid species (D. kaki, in particular), there is an insertion of a retroelement, named Kali, in the promoter region of the OGI gene that prevents the synthesis of small RNA and promotes the development of monoecy.
2.2.4. Pistachio
2.2.5. Hippophae
he sea buckthorn karyotype consists of 24 chromosomes (22 autosomes and a pair of X/Y sex chromosomes) [163]. In the genome (about 2.6 pg in size), researchers have observed a uniquely large number of satellites that may turn out to be good cytogenetic markers [163,164,165]. The development of molecular markers to determine the sex of sea buckthorn in the early stages of cultivation has been quite successful, but the experiments have shown the unstable operation of these markers, which may be due to sea buckthorn’s polymorphism [166,167,168]. Morphologically, male and female plants can only be distinguished by their generative buds after flowering age has been reached. At the same time, mainly female plants are needed in agricultural production. Additionally, breeding is carried out independently for male pollinating plants and female plants [169]. These facts demonstrate the need for a deeper study of the genetic determination of the sex in sea buckthorn.
2.2.6. Humulus
Humulus lupulus L. is a dioecious woody vine of the Cannabaceae family that is of great importance in brewing. Hops are grown for the female buds, which contain substances that create the characteristic taste, aroma and bitterness of beer. Due to the popularity of this drink, hops are cultivated on every continent except Antarctica. The hop karyotype consists of 18 autosomes and a pair of sex chromosomes (XX/XY). The Y chromosome is the smallest in the karyotype, which may indicate its degeneration and evolutionary antiquity [172,173,174]. In commercial cultivation, the female varieties are vegetatively propagated. A very important task is to prevent the pollination of inflorescences, since setting seeds spoils the taste of beer [175]. Hop is a wind-pollinated crop with light pollen, so just one male plant in the area can cause a lot of economic damage [176]. To prevent pollination, all male plants in a nursery and nearby wild populations must be eradicated. As a part of this procedure, molecular genetic markers are used to identify sex in the early stages of ontogenesis [38,177,178]. In hop production, seedless triploid varieties are also cultivated. They are more productive and have a high growth rate [179,180].
2.2.7. Date Palm
2.2.8. Myristica
2.2.9. Actinidia
Morphologically, the flowers look bisexual, but female plants form flowers with sterile pollen while male plants have an underdeveloped and non-functioning pistil [197]. However, some species sometimes have bisexual flowers. Hermaphroditic flowers have been observed in A. arguta, A. chinensis, A. deliciosa, and A. eriantha Benth. [196]. Kiwifruit has only been cultivated for just over 100 years, but it has achieved great popularity around the world, and its production is about 4 million tons per year [46]. Polyploidy is widespread in species of the Actinidia genus [198]. Chromosomal numbers range from 2n = 58 to 2n = 174 (x = 29) [199]. Polyploidy does not appear to affect sex in kiwifruit. Male plants have at least one Y chromosome in the karyotype, and female plants have XX [200,201]. The sex chromosomes are small and homomorphic, with a small SDR [41,202]. The mechanism of sex determination has been widely studied. This mechanism is based on two genes (the Shy Girl gene (SyGI), the dominant suppressor of carpel development, and the Friendly Boy gene (FrBy), expressed in tapetum cells) [15]. Currently, the kiwifruit genome is actively being studied, and genetic maps and molecular markers are being created to aid the breeding of this crop [203,204,205,206]. However, hermaphroditism can be determined by additional genetic factors, which creates difficulties for sex determination based on molecular markers [206].
2.2.10. Ilex
2.2.11. Chinese Bayberry
Chinese bayberry (Morella rubra Lour.) is widely cultivated in China for fruits. It is the only edible species of the Myricaceae family [219]. The chromosome number is 2n = 16, and the heterogametic sex is presumably female (ZW chromosome system) [220]. A comparison of the sequenced genomes of male and female plants revealed a small region (59 kb) on chromosome 8 that is specific to female plants [42]. The differential expression of some genes, presumably associated with the development of sex in this species, was also shown [221].
3. Conclusions
This entry is adapted from the peer-reviewed paper 10.3390/agronomy13020540