Your browser does not fully support modern features. Please upgrade for a smoother experience.
Please note this is an old version of this entry, which may differ significantly from the current revision.
Subjects:
Plant Sciences
Environmental metal pollution is a common problem threatening sustainable and safe crop production. Heavy metals (HMs) cause toxicity by targeting key molecules and life processes in plant cells. Plants counteract excess metals in the environment by enhancing defense responses, such as metal chelation, isolation to vacuoles, regulating metal intake through transporters, and strengthening antioxidant mechanisms. In recent years, microRNAs (miRNAs), as a small non-coding RNA, have become the central regulator of a variety of abiotic stresses, including HMs.
- heavy metals
- miRNA
- toxicity response
1. Introduction
The rapid growth of industrialization and the large-scale use of chemical fertilizers and pesticides have led to the continuous increase in heavy metal content in the soil. In plants, copper (Cu), zinc (Zn), iron (Fe), and manganese (Mn) are trace elements necessary for plant development and growth, but excessive accumulation can cause cell damage [1]. Some other metals such as cadmium (Cd), chromium (Cr), lead (Pb), aluminum (Al), arsenic (As), and mercury (Hg), as non-essential elements, are toxic even at low concentrations [2]. Studies have reported that heavy metal stress can inhibit activity of antioxidant enzyme systems and differential expression of a large number of proteins in plants, weakening photosynthesis, accompanied by a series of phenotypes such as suppressed root development and leaf senescence or even necrosis [3]. In order to avoid the destructive consequences of heavy metal toxicity, plants have developed corresponding coping mechanisms to resist heavy metal stress [3]. Plants can reduce heavy metal concentrations in their bodies by limiting the uptake of heavy metal ions and stimulating metal efflux, as well as by complexation of metal ligands such as glutathione (GSH), metallothionein (MT), and phytochelatins (PCs) [4,5,6]. In addition, the antioxidant defense mechanism will also be activated to reduce elevated reactive oxygen species (ROS) levels, thereby reducing oxidative damage [7,8]. A large number of studies have shown that gene expression plays a very important role in regulating the tolerance of HMs or individually regulating various stress response genes to form a gene network. Some functional group genes encode metabolites such as amines, alcohols, and sugars, which also play a crucial role in heavy metal stress tolerance [9,10].
With the discovery of small RNAs, there is growing interest in the importance of post-transcriptional gene regulation of microRNAs (miRNAs) in plant development and responses to environmental stresses. miRNAs are an extensive class of small noncoding (19~24 nt) RNAs molecules [11]. In plants, mature miRNAs are produced through a multistep process including the transcription, precursor processing, methylation, and assembly of miRNA-induced silencing complex (miRISC) [12]. Then, the mature miRNAs lead the RISC to target the complementary mRNAs, which play a vital post-transcriptional regulatory role in gene expression by target mRNA cleavage or translational inhibition [13]. In recent years, a crescendo of miRNA studies has demonstrated that miRNAs play important roles in tissue development and differentiation, phytohormones signaling, secondary metabolite production, and biotic and abiotic stress [14,15]. miRNAs affect multiple processes of plant growth, development, and response to stress by up-regulating or down-regulating their expression [16,17]. miR165/166 and miR394 have been shown to be involved in shoot apical meristem (SAM) development, including the direct post-transcriptional regulation of key SAM-related genes, which in turn maintains SAM development [18,19]. In addition, studies have shown that miR319 has a conserved regulatory function in leaf development. For example, ectopic upregulation of miR319 resulted in dramatic changes in tomato leaf size and shape [20]. CUC1 and CUC2 mRNAs accumulate in the axil of leaf primordia and play a key role in the establishment of axillary bud meristems. The regulatory mechanism of miR164-CUC1/CUC2 may be related to LAS-mediated initiation of axillary bud meristems [21]. In addition, miRNAs play important roles in plant root development. miR160 acts as a key controller and cleaves ARF10 and ARF16 transcripts during plant root cap formation [22]. In contrast, miR167 plays an active role in adventitious root formation, while miR156 and miR172 are well-studied miRNAs involved in floral control [23,24]. Overexpression of miR172 can promote flowering time in both monocotyledonous and dicotyledonous plants. In contrast, the expression level of miR156 decreased gradually from sowing to flowering, while upregulation of miR156 resulted in delayed flowering transition. These findings suggest that miRNAs play regulatory roles in different developmental transitions by mediating specific signaling pathways. In different models and crop plants (such as Arabidopsis, wheat, rice, maize, and barley), miRNAs regulate gene expression during different stress responses (drought, heat, salinity, cold, nutrition, and pathogens). In addition to plant growth and development, the role of conserved miRNA target modules is also critical for conferring stress tolerance through integration into metabolic pathways [25]. Studies have confirmed that miR160-ARF, miR156-SPL, miR159-MYB33, miR164-NAC, miR172-AP2, miR394-LCR, miR396-GRF, and miR398-CSD modules play important regulatory roles in different stress environments to mitigate the effects of adverse reactions [12]. For example, in Arabidopsis, increased expression of miR398 enhances plant heat tolerance by negatively regulating the expression of its targets CSD1, CSD2, and the copper chaperone (CCD) of CSD [26]. The highly conserved miR394-LCR module is involved in plant responses to cold stress [27]. The expression of auxin-responsive factors ARF10, ARF16, and ARF17, mediated by miR160 and miR167, resulted in enhanced salinity tolerance of cotton under high salt stress. Overexpression of osa-miR319a exhibited higher tolerance to drought and salt stress by regulating the TCP transcription factor [28].
Increasing evidence has also revealed that miRNA-mediated gene regulation plays a significant role in heavy metal regulatory networks. In addition, high-throughput genome-wide expression profiling has greatly improved our current understanding of the key involvement of miRNA in the toxic response of plant HMs and its targets. A substantial number of heavy-metal-responsive miRNAs have been identified in the Oryza sativa L., Zea mays L., Medicago truncatula L., Hordeum vulgare L., Vitis vinifera L., Brassica juncea L., and many other plants (Table 1) [6,29,30]. In different plant species, many miRNAs have significant differential expression of different HM such as Cd, Hg, Al, As, and Cr. Through the analysis of metal-regulated miRNA target genes, it has also been identified that many miRNAs are involved in the response of plants to HM. The target genes for various processes, including metal absorption and transport, sulfate distribution and assimilation, protein folding, antioxidant systems, and plant hormone signal transduction processes [11]. We searched the Web of Science core collection database for articles related to miRNA and heavy metals such as Cu, Cd, Hg, Al, As, and Cr in the past 10 years, and a total of 568 papers were retrieved. The research fields are mainly distributed in three aspects: Life Sciences Biomedicine, Science Technology, and Physical Sciences. Then, bibliometric analysis was performed using VOS software [31], and four clusters were formed with miRNAs, stress, cadmium, and strategy as the research centers (Figure 1). These keywords are categorized by publication year to deepen the analysis. The VOSviewer software represents the year with the highest number of publications between 2016 and 2019. However, articles from 2011 to 2022 were included in this analysis. The blue circles represent topics that have been intensively studied in the past decade, such as stress response, miRNA, target gene, etc. The results of the two co-occurrence maps showed that miRNAs and stress were most closely related, suggesting that the involvement of miRNAs in stress response is the current research hotspot.
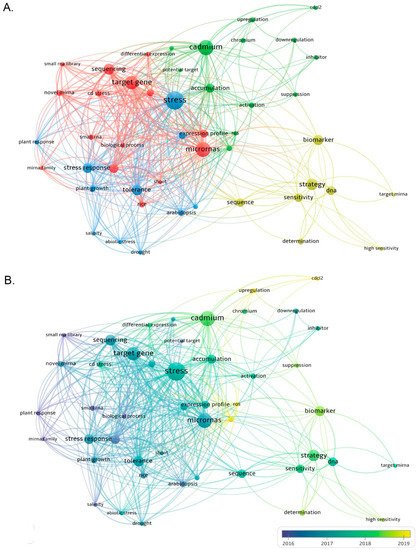
Figure 1. Keyword co-occurrence graph analysis using VOSviewer software. (A) A total of 568 relevant articles were searched from academic papers over the past 10 years using the keywords miRNA, Cu, Cd, Hg, Al, As and Cr, four research centers focusing on miRNA, stress, cadmium, and strategy were formed. (B) Different colors correspond to the year in which the keywords appeared on average, and keywords with blue color presented earlier than those with yellow.
Table 1. List of miRNAs along with their target function during heavy metal toxicity tolerance.
| miRNA | Plant Species | Stress | Target Genes | Target Function | Reference |
|---|---|---|---|---|---|
| miR156 | A. thaliana, V. Vinifera, O. sativa, Nicotiana tabacum L., Miscanthus sinensis A. |
Cu, Cd, Cr, Al | SPL7 | Regulate Cu homeostasis, decrease endogenous ROS | [32,33,34,35,36,37] |
| miR159 | Brassica napus L., O. sativa, N. tabacum, M. truncatula, Saccharum spp | Cd, As, Cr, Al | ABC transporter protein, OPT1 | Regulate metal transporters | [35,36,38,39,40,41] |
| miR160 | A. thaliana, O. sativa, Saccharum spp | Cr, Al | ARF | Regulate auxin signal | [41,42] |
| miR164 | A. thaliana, O. sativa, M. sinensis, Saccharum spp |
As, Cr, Al, Hg | NAC, CUP trancription factors | Signaling pathway, root development, response to oxidative stress | [41,43,44] |
| miR166 | O. sativa, N. tabacum | Cd, Cr | Homeodomain containing protein 4 | Reduced Cd translocation from roots to shoots and Cd accumulation in seeds, decrease Cd-induced oxidative stress | [35,36,45] |
| miR167 | O. sativa, B. juncea, N. tabacum | Cd, As | NRAMP1b metal transporter protein | Metal uptake and translocation | [36,46,47] |
| miR168 | O. sativa, | Cd | LOX1 | Promote JA synthesis | [48] |
| miR169 | M. truncatula, O. satva, N. tabaccum, B. juncea | Cr, Cd, Al, As | CCAAT-binding (TF) | ? | [35,38,44,49] |
| miR171 | M. truncatula, B. Juncea, O. sativa, N. tabacum |
Hg, As, Cr, Cd | SCL (TF) | Shoot branching, signaling pathway |
[2,35,36,50] |
| miR192 | O. sativa | Cd | ABC gene | Seed germination of rice under Cd stress | [51] |
| miR268 | O. sativa | Cd | NRAMP3 | Inhibited rice seedling growth under Cd-stress treatment | [52] |
| miR319 | M. truncatula, B. juncea, M. truncatula | Hg, As, Al | TCP (TF), cyclin | Leaf morphogenesis, cell differentiation, embryonic development, cell division |
[47,50,53] |
| miR390 | O. sativa, M. truncatula, Glycine max L. | As, Al | tasi-RNA | Plant development | [40,54,55] |
| miR393 | A. thaliana, O. sativa, M. truncatula, Saccharum spp, H. vulgare | Cd, Hg, Al | TIR1/AFBs (F -box auxin receptors) and bHLH (TF) | Regulate auxin signaling | [41,50,56,57,58] |
| miR395 | A. thaliana, B. napus, Brassica parachinensis L. | Cd, As, Al | SLIM1, SULTR2;1, APS | Sulfur assimilation, response to cadmium ion, sulfate transport |
[43,59,60,61,62] |
| miR396 | O. sativa, N. tabacum, M. sinensis, M. truncatula, G. max | As, Cr, Al | GRF (TF) | Translation, leaf development | [36,37,40,43,55] |
| miR397 | A. thaliana, O. sativa, Populus trichocarpa T. | Cu, As, Cr | Laccase | Regulate inter-tissue lignification and secondary cell wall thickness, activities of PPO, SOD, and POD | [35,59,63,64,65,66] |
| miR398 | A. thaliana, O. sativa, P. trichocarpa | Cu, Cd | CSD1, CSD2, CCS1 and COX5b-1 | SODs relive oxidative stress | [34,65,67,68] |
| miR399 | O. sativa, N. tabacum | As | ? | ? | [36,43] |
| miR408 | A. thaliana, O. sativa, P. trichocarpa, O. sava | Cu, Cr, As | Laccase, plantacyanin transcripts, SPL7, HY5 | Regulate plastocyanin (PC) content | [63] [35,43,65,69,70] |
| miR444 | O. sativa | Cr | ? | ? | [35] |
| miR528 | O. sativa, A. thaliana, Z. mays | As, Al, Cd | MAX2 gene and l-resistant blo, LACod acid oxidase, MATE, LAC | Signal transduction, regulation of cell cycle, plant development, ascorbate metabolism, miRNA processing, control of cellular-free auxin levels | [43,54,70,71,72,73] |
| miR529 | O. sativa, M. truncatula |
As, Cd | Apetala2-like (TF), squamosa promoter binding protein-like (TF) | Signaling pathway, plant development | [43,50] |
| miR808 | O. sativa, | Al | Rockweed glycosyltransferase | ? | [74] |
| miR854 | B. juncea | As | Putative serine acetyl transferase (SAT) | Synthesis of O-acetylserine | [47] |
| miR857 | A. thaliana, | Cu | Laccase (LAC7) | ? | [33] |
| miR1318 | O. sativa | As | Calcium-binding proteins or Ca2+ ATPase | Signaling | [70] |
| miR1444 | P. trichocarpa | Cu | Cu-containing proteins, polyphenol oxidases (PPOs) | ? | [65] |
| miR1535b | G. max | Cd | Glyma07g38620.1 | Responsible for initial step of isopentenyl transferase |
[75] |
In recent years, research on miRNA and HM stress has been increasing, and, with the rapid development of advanced technologies such as next-generation sequencing (NGS), a number of known and unknown miRNAs in response to metal stress are discovered through whole genome sequencing and miRNA sequencing. However, studies on the roles of different miRNAs in HMs signal transduction in plants and their targets are still in their infancy.
2. Response of miRNAs to Cu Toxicity
Cu is a trace element essential for plant growth and development. In animals, plants, and microorganisms, it is usually found in the form of Cu ions or cuprein. As an important cofactor of protein, Cu is a component of polyphenol oxidase, superoxide dismutase, laccase, cytochrome oxidase, and other enzymes, and is involved in important physiological processes such as photosynthesis, respiratory metabolism, and oxidative stress [76,77]. Cu is also a component of plastocyanin and participates in the electron transfer process of photosynthesis. However, the deficiency or excessive accumulation of Cu can cause damage to plant growth. Cu deficiency can lead to blue-green, wrinkled, distorted, or necrotic leaves, dwarf plants, slow growth, and reduced yields [78]. In contrast, excess Cu induces rapid synthesis of oxidation anions (O2−), hydroxyl radicals (OH), hydrogen peroxide (H2O2), singlet oxygen (1O2), and ROS in plant cells [79]. This reduces the strength of the cell membrane, leading to the toxic effects of Cu2+ infiltration into the cells. Excess Cu also inactivates chloroplast enzyme activity, accelerates chloroplast decomposition, inhibits chlorophyll synthesis, or rapidly compounds and destroys chlorophyll in plant cells. Additionally, Cu stress causes the electron-transport chain to be blocked, affecting plant photosynthesis [80]. Cu toxicity also affects the normal uptake of other mineral nutrients by plant roots. Cu stress disrupts the structure of protoplasts and affects their function, changing the permeability of cell membranes to increase, leading to the leakage of various ions from the membrane, and disrupting ionic equilibrium and a corresponding decrease in nutrient content [81]. To maintain the correct concentration of Cu2+ in cells, plants develop an important regulatory network in Cu uptake, distribution, and molecular responses to frequent Cu changes. The involvement of a group of miRNAs in this network appears to be particularly important, as they regulate many functionally distinct Cu proteins, including laccases, plastocyanins (PC), Cu/Zn superoxide dismutase, and polyphenol oxidase. In recent years, many studies have found that the miRNAs involved in copper stress response mainly include miR397, miR398, miR408, miR857, and miR1444, among which miR397, miR398, and miR408 are conserved in Arabidopsis and rice [32,67].
miR398 is a highly conserved miRNA in terrestrial plants, and there are three members of the miR398 family in Arabidopsis (miR398a, miR398b, miR398c) [32,67]. The sequence of miR398b and miR398c are identical, and miR398a differs from them by only one nucleotide at the 3′ end [67]. Compared with miR398b and miR398c, the promoter sequence of miR398a does not contain the GTAC sequence, which can explain the low expression of miR398a and the slow response to Cu deficiency [67]. In addition, miR398 regulates three genes through transcript cleavage and translational inhibition, including the cytosolic (CSD1) and plastidic (CSD2) genes in the Cu/Zn superoxide dismutase gene family, the copper chaperone for the superoxide dismutase (CCS1) gene [67]. The CSD1 and CSD2 genes encode closely related Cu/Zn superoxide dismutases, which detoxify superoxide free radicals. CCS1 is an intracellular Cu transporter protein with the function to transport Cu ions to the CSD through protein-to-protein interactions, which in turn activates the CSD. Under high Cu stress, the expression of miR398 suppressed, resulting in an increase in CSD1 and CSD2 expression, which relieves the threat of ROS due to the increased Cu content [67]. In the absence of Cu, the expression of miR398 would be induced, and the transcription levels of its target genes CSD1 and CSD2 would decrease, while iron superoxide dismutase (FeSOD) will be up-regulated to increase the Cu availability of other important Cu proteins (such as plastocyanin) [67]. miR398 plays a key role in the regulation of Cu homeostasis by regulating the non-essential Cu protein CSD, when Cu is lacking or excessive.
Meanwhile, miR397, miR408, and miR857 are also involved in regulating the abundance of other Cu proteins in Arabidopsis, especially the effectiveness of laccase and the secreted protein plantacyanin in response to Cu [33]. As a Cu-containing oxidase, laccase can promote the synthesis of lignin from lignin monomer in plants and promote the lignification process of plants [82]. miR397 is a key regulator of Cu homeostasis in plants such as A. thaliana, P. trichocarpa, and V. vinifera. Under low Cu stress, the expression of miR397 was up-regulated, the synthesis of laccase or plastocyanin was inhibited, and the accumulation of Cu increased, which was beneficial to maintain the homeostasis of Cu [63,64,83]. In tomato (Solanum lycopersicum M.), after overexpression of miR397a, its target gene LeLACmiR397a was down-regulated, which caused a decrease in the activities of polyphenol oxidase (PPO), superoxide dismutase (SOD), and peroxidase (POD). In addition, miR397 is also related to the yield of crops. Compared with wild-type plants, transgenic rice that overexpresses miR397 has larger seeds, increased inflorescence numbers, and increased yields [59]. Studies have shown that miR408 can respond to abiotic stress and maintain Cu homeostasis in plants [69]. A study showed that the miR408 could bind to the 5`-UTR region of LAC1, LAC12, and LAC13, and regulated the expression of these genes to maintain Cu homeostasis in plants [63,69]. It was demonstrated that transgenic strains with simultaneous inhibition of the functions of three conserved Cu-miRNAs (miR397, miR398, and miR408) showed the reduced accumulation of Cu-miRNAs, the increased accumulation of transcripts encoding Cu proteins, and that photosynthesis and growth of transgenic plants were affected under low Cu conditions, which may be associated with defective accumulation of chloroplast plastocyanin [84]. Interestingly, miR1444 is currently only found in P. trichocarpa and appears to be a Populus-specific miRNA. It has also been shown to regulate a group of Cu-containing proteins: polyphenol oxidases (PPOs) [65].
The study found that the promoter regions of these several miRNAs all have Cu-response elements (CuRE) as the role sites of Cu regulation, which is composed of repeated GTAC sequences and is the only Cu-responsive element identified to date [85]. SQUAMOSA promoter-binding protein-like-7 (SPL7) is a key transcription factor that is sensitive to low Cu and plays a transcriptional regulatory role by binding to GTAC sequences [68]. In Cu deficiency, SPL7 can activate the transcription of miR397, miR398, miR408, and miR857 genes, which inhibit the expression of copper-containing protein genes such as laccase and Cu-Zn superoxide dismutase, thereby regulating Cu in the case of Cu-deficiency allocation [68]. When Cu accumulates, SPL7 becomes inactive and the expression of miRNAs in plants are inhibited, which increases the transcription level of target genes and improves the availability of Cu proteins, thus alleviating the oxidative damage caused by copper accumulation (Figure 2).
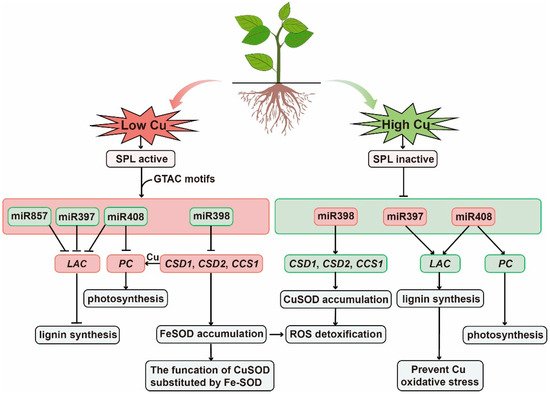
Figure 2. Schematic representation of the mode of action of miRNAs in response to Cu stress. Green boxes indicate relevant miRNAs and pathways involved in low Cu stress. SPL7 transcription factors are active and regulate miR397, miR398, miR408, and miR857, which regulate genes encoding Cu-containing proteins, including Cu/Zn superoxide dismutase (CSD), CCS1, laccase (LAC), and phycocyanin (PLC), thereby conserving Cu as essential Cu protein (such as plastocyanin). Red boxes indicate that under high Cu stress, SPL7 is inactivated, miR398 is down-regulated, and the expression of target genes CSD1, CSD2, and CCS1 is up-regulated, mitigating the threat of ROS from increased Cu content and increased Cu protein accumulation.
With the development of transcriptome and small RNA sequencing technology, a large number of studies will utilize this method to identify miRNAs in plants. In 2015, under the control and Cu stress conditions, the conservative and non-conservative miRNA and other short RNA were identified in Paeonia ostia T. [86]. A total of 102 known plant miRNAs were identified and combined with transcriptome sequencing data, while 34 new potential miRNAs were identified under the same conditions. It was also found that 12 conservative miRNA and 18 new miRNAs changed significantly under Cu stress. Jiu et al. (2019) used high-throughput sequencing to determine the miRNAs and target genes of grapevine (V. vinifera) plants responsive to Cu stress [87]. Among them, 100 known and 47 newly discovered miRNAs were differentially expressed under Cu stress. The target prediction of miRNA shows that miRNA may regulate transcription factors such as AP2, SBP, NAC, MYB, and ARF under Cu stress. They also found that miR156 targets SPL7 and may be involved in the regulation of Cu homeostasis in grapes (V. vinifera). In addition, a total of 65 known miRNAs and 78 new miRNAs predicted to mature in mulberry were identified [30]. In total, 40 miRNAs were differentially expressed under Cu stress, of which 27 miRNAs up-regulated genes and 13 miRNAs down-regulated genes. Utilizing high-throughput sequencing technology, a large number of unknown miRNAs have been found to be involved in the Cu-stress-response process. How these miRNAs regulate the distribution of Cu in plants, enabling plants to coordinate the expression and development of Cu proteins, requires further studies to explore.
This entry is adapted from the peer-reviewed paper 10.3390/ijms23158642
This entry is offline, you can click here to edit this entry!