Pollination is essential to maintain ecosystem balance and agricultural production. Domesticated bee pollination, which is easy to feed and manage, and mechanized pollination, which is not restricted by the environment, are considered the main technical means to alleviate the “pollinating insect crisis”. The selection of pollination method should be based on the physiological characteristics of crops and the actual environmental conditions of natural pollination. The pollination ability of bees is closely related to the status of the bees. Maintaining the pollination ability of bees in a reasonable range is the goal of bee pollination services. Colony state control technology needs to develop in two directions. First, there is a need to develop colony state monitoring technology based on multi-feature information fusion and to explore the self-regulation mechanisms of the colony in response to various factors. Second, based on these self-regulation mechanisms, there is a need to develop a low-cost and non-invasive bee colony state and pollination capacity estimation model, monitoring technology, and equipment based on single feature information. The goals of mechanized pollination are “efficiency” and “precision”. Mechanized pollination technology needs to be developed in two directions. First, the mechanisms of pollen abscission, transport, and sedimentation in different crops and mechanized pollination conditions should be explored. Second, research and development of efficient and accurate pollination equipment and technology based on the integration of multiple technologies such as pneumatic assistance, auxiliaries, static electricity, target, variables, and navigation, are needed.
- bee pollination
- mechanized pollination
- crops
- efficient pollination technology
1. Introduction
2. Pollination Methods for Different Crops
2.1. Basic Structural Features of Flowers
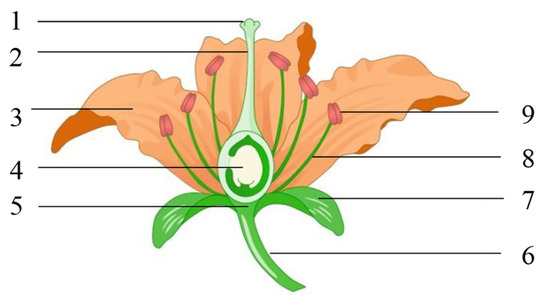
2.2. Self-Pollination and Cross-Pollination
2.3. Differences in Pollination Methods of Different Crops
3. Status of Efficient Bee Pollination Technology
3.1. Principle of Efficient Pollination of Bees
Bee pollination is a “bonus” of bees’ out-foraging behavior. In the long-term coevolution of bees and plants, bees have formed numerous characteristics suitable for plant pollination, including being easily attracted by bright colored entomophilous flowers that emit an aroma and secrete nectar; in addition, bees have identifying memory and pollination activity focused on specific species over a long period, high-frequency vibration generated by the wings during foraging that causes pollen to fall off, morphological structures such as villi and pollen baskets that easily adhere to pollen, social group structure, and numerous groups [21].
3.2. Breeding of High-Quality Pollinating Bee Varieties
3.3. Climate and Environment Control Technology Outside the Hive
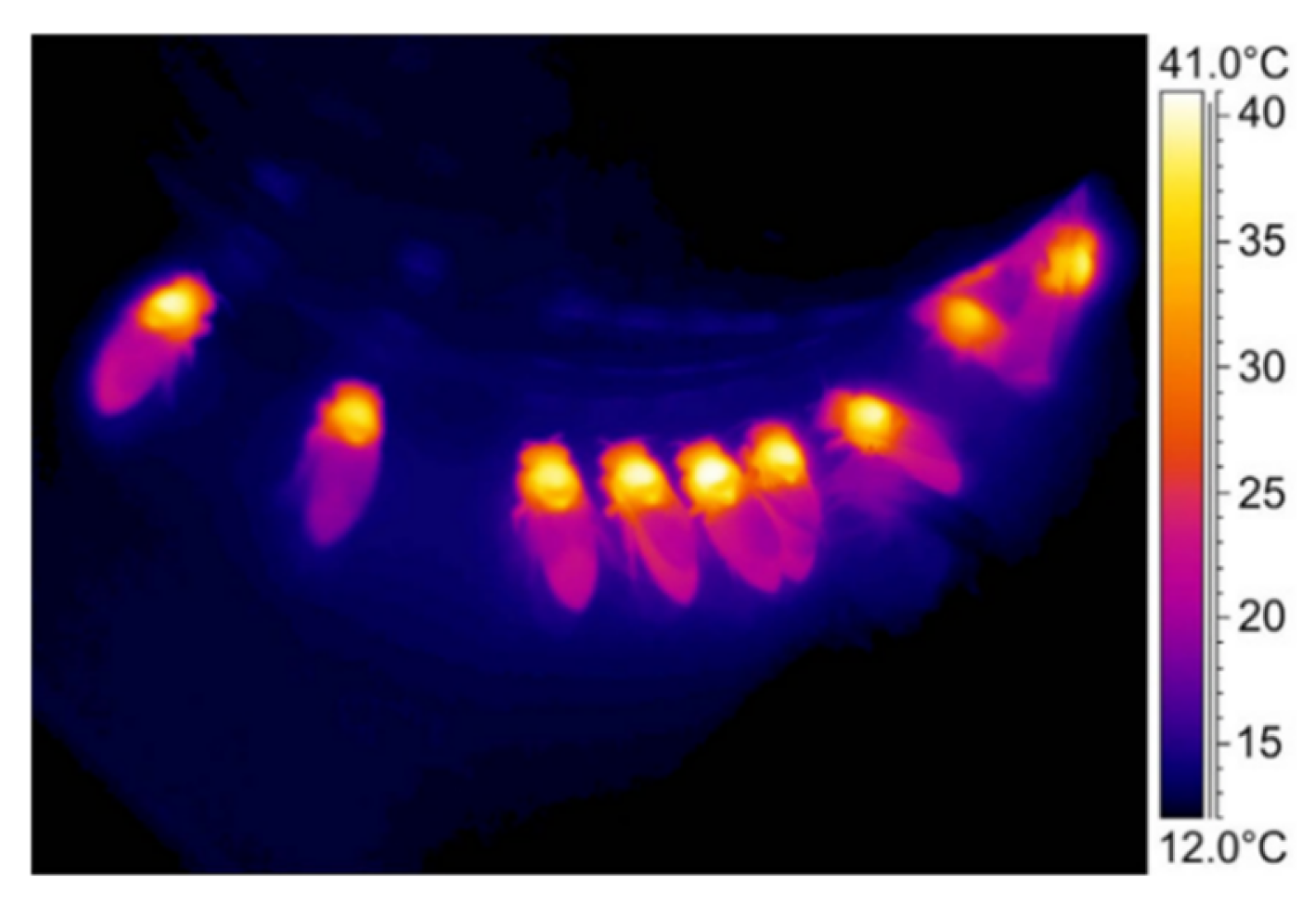
3.4. Temperature and Humidity Control Technology in the Beehive
Healthy breeding of queen bees, drones, and worker bees is the basis for the health, strength, and stability of the colony. The beehive provides a breeding and living place that shields the bee colony from light, rain, and wind. Temperature and humidity in the beehive are the most critical factors that affect colony reproduction and are directly affected by the climate outside the beehive.
3.4.1. Influence of Temperature and Humidity on Beehive Colony Reproduction
3.4.2. Colony Self-Regulation Mechanism of Beehive Temperature and Humidity
Honeybee antennae are multimodal sensory organs that can sense various information, such as temperature, humidity, smell, taste, and mechanical stimuli, among others. Their temperature sensitivity is 0.25 °C. Honeybees receive real-time information on temperature, humidity, and spleen temperature in the hive through their antennae [43][44][45]. When the temperature, humidity, and temperature of the spleen in the hive are beyond the reasonable range for breeding and life, the bee colony takes measures to modify it. When the temperature in the hive or spleen is low, thermogenic bees raise the temperature of their thoracic cavity and attach the breast to the wax cover of the sealing lid, transferring this heat to maintain the temperature in the hive and spleen. Non-thermogenic bees gather and squeeze the cell comb to increase the sealing performance of the hive and reduce heat loss [46][47]. When the temperature in the beehive or spleen is high, the bee colony increases airflow between the inside of the beehive and the outside world through ordered directional fans to discharge the overheated air in the beehive; furthermore, the bee colony is also scattered in the beehive, further improving ventilation and heat dissipation [48].
3.4.3. Temperature and Humidity Monitoring Technology in Beehives
The existing temperature and humidity control in the beehive mainly relies on the self-regulating ability of the bee colony. A healthy, strong, and stable bee colony can maintain the temperature and humidity in the beehive within a reasonable range to ensure the breeding and life of the bee colony. A typical winter colony of 17,500 bees can survive for over 300 h even when the temperature outside the hive is as low as −25 °C [49]. However, when the state of the bee colony changes because of, for example, the occurrence of disease and insect pests, separation of bees, escape, or changes in the temperature and humidity inside the beehive caused by external changes that exceed the ability of the existing bee colony to adjust, the survival of the entire bee colony is seriously threatened.
3.5. Bee Colony State Management and Control Technology
3.5.1. Image Monitoring Technology in Beehives
3.5.2. Sound and Vibration Monitoring Technology in Beehives
The sound and vibration emitted by bees are an essential part of the communication mechanism of the bee colony. The temperature and humidity regulation of the colony, group activities such as going out for food, separating bees, escaping, losing the king, and disease can all be reflected by sound and vibration information.
3.5.3. Bee Colony Weight Monitoring Technology
3.5.4. Bee Colony Monitoring Technology in the Hive Gate Area
3.5.5. Bee Colony Monitoring Technology Based on Multi-Feature Information Fusion
4. Status of Mechanized Pollination Technology
4.1. Principle of Mechanized Pollination
4.2. Mechanized Pollination Technology and Equipment for Field Crops
4.3. Mechanized Pollination Technology and Equipment for Orchards
4.4. Mechanized Pollination Technology and Equipment for Greenhouses
5. Conclusions
This entry is adapted from the peer-reviewed paper 10.3390/agronomy12112872
References
- Ollerton, J.; Winfree, R.; Tarrant, S. How many flowering plants are pollinated by animals? Oikos 2011, 120, 321–326.
- Klein, A.-M.; Vaissiere, B.E.; Cane, J.H.; Steffan-Dewenter, I.; Cunningham, S.A.; Kremen, C.; Tscharntke, T. Importance of pollinators in changing landscapes for world crops. Proc. R. Soc. B-Biol. Sci. 2007, 274, 303–313.
- Gallai, N.; Salles, J.-M.; Settele, J.; Vaissiere, B.E. Economic valuation of the vulnerability of world agriculture confronted with pollinator decline. Ecol. Econ. 2009, 68, 810–821.
- Kjøhl, M.; Nielsen, A.; Stenseth, N.C. Potential Effects of Climate Change on Crop Pollination; Food and Agriculture Organization of the United Nations: Rome, Italy, 2011.
- Connolly, C. The risk of insecticides to pollinating insects. Commun. Integr. Biol. 2013, 6, e25074.
- Maes, J.; Hauck, J.; Paracchini, M.L.; Ratamaki, O.; Hutchins, M.; Termansen, M.; Furman, E.; Perez-Soba, M.; Braat, L.; Bidoglio, G. Mainstreaming ecosystem services into EU policy. Curr. Opin. Environ. Sustain. 2013, 5, 128–134.
- FitzPatrick, Ú.; Stout, J.; Bertrand, C.; Bradley, K.; Clabby, G.; Keena, C.; Walsh, J. All-Ireland Pollinator Plan 2015–2020; National Biodiversity Data Centre: Waterford, Ireland, 2015.
- Hung, K.-L.J.; Kingston, J.M.; Albrecht, M.; Holway, D.A.; Kohn, J.R. The worldwide importance of honey bees as pollinators in natural habitats. Proc. R. Soc. B-Biol. Sci. 2018, 285, 20172140.
- Binns, C. Robotic Insects Could Pollinate Flowers and Find Disaster Victims Popular Science USA. 2009. Available online: https://www.popsci.com/technology/article/2009-12/flight-robobee/ (accessed on 10 November 2022).
- Leins, P.; Erbar, C. Flower and fruit. Hoppea 2010, 71, 354–355.
- Brown, A.H.D.; Zohary, D.; Nevo, E. Outcrossing rates and heterozygosity in natural populations of Hordeum spontaneum Koch in Israel. Heredity 1978, 41, 49–62.
- Vuletin Selak, G.; Perica, S.; Ban Goreta, S.; Radunic, M.; Poljak, M. Reproductive Success after Self-pollination and Cross-pollination of Olive Cultivars in Croatia. HortScience 2011, 46, 186–191.
- Chavez, J.D.; Lyrene, M.P. Effects of Self-pollination and Cross-pollination of Vaccinium darrowii (Ericaceae) and Other Low-chill Blueberries. HortScience 2009, 44, 1538–1541.
- Lloyd, D.G.; Webb, C.J. The avoidance of interference between the presentation of pollen and stigmas in angiosperms I. Dichogamy. N. Z. J. Bot. 2011, 24, 135–162.
- Webb, C.J.; Lloyd, D.G. The avoidance of interference between the presentation of pollen and stigmas in angiosperms II. Herkogamy. N. Z. J. Bot. 1986, 24, 163–178.
- Trelease, W. The Heterogony of Oxalis violacea. Am. Nat. 1882, 16, 13–19.
- East, E.M. The distribution of self-sterility in the flowering plants. Proc. Am. Philos. Soc. 1940, 82, 449–518.
- Lord, E.M. Cleistogamy: A Tool for the Study of Floral Morphogenesis, Function and Evolution. Bot. Rev. 1981, 47, 421–449.
- Kumar, S.; Rao, M.; Gupta, N.C. Breeding Strategies of Self Pollinated Crop with Special Emphasis on Hybrid Rice: Present and Future Perspectives. Res. Rev. J. Agric. Sci. Technol. 2014, 3, 2349–3682.
- Dicenta, F.; Ortega, E.; Canovas, J.A.; Egea, J. Self-pollination vs. cross-pollination in almond: Pollen tube growth, fruit set and fruit characteristics. Plant Breed. 2002, 121, 163–167.
- Winston, M.L. The Biology of the Honey Bee; Harvard University Press: Cambridge, MA, USA, 1991.
- Osterman, J.; Aizen, M.A.; Biesmeijer, J.C.; Bosch, J.; Howlett, B.G.; Inouye, D.W.; Jung, C.; Martins, D.J.; Medel, R.; Pauw, A. Global trends in the number and diversity of managed pollinator species. Agric. Ecosyst. Environ. 2021, 322, 107653.
- Zhigang, L.; Hongsong, R.; Guangyu, F.; Aihemaiti, M.; Hu Xidan, M.; Kurban, A.; Ruihua, W.; Hongmei, G.; Jian, W.; Haifeng, L. Effect of Bumblebee Pollination on Yield and Quality of Greenhouse Tomato; Jiangsu Academy of Agricultural Sciences: Nanjing, China, 2021.
- Hongdong, W.; Shuang, H.; Bing, H.; Mingfei, W.; Guanxiong, Z.; Donggang, L. Research progress of bumblebee pollination technology in protected agriculture. Yangtze River Veg. 2022, 8, 34–37.
- Joshi, N.C.; Joshi, P.C. Foraging Behaviour of Apis spp. on Apple Flowers in a Subtropical Environment. N. Y. Sci. J. 2010, 3, 71–76.
- Blazyte-Cereskiene, L.; Vaitkeviciene, G.; Venskutonyte, S.; Buda, V. Honey bee foraging in spring oilseed rape crops under high ambient temperature conditions. Zemdirb. Agric. 2010, 97, 61–70.
- Abrol, D.P. Diversity of pollinating insects visiting litchi flowers (Litchi chinensis Sonn.) and path analysis of environmental factors influencing foraging behaviour of four honeybee species. J. Apic. Res. 2006, 45, 180–187.
- Peat, J.; Goulson, D. Effects of experience and weather on foraging rate and pollen versus nectar collection in the bumblebee, Bombus terrestris. Behav. Ecol. Sociobiol. 2005, 58, 152–156.
- Kovac, H.; Stabentheiner, A. Thermoregulation of foraging honeybees on flowering plants: Seasonal variability and influence of radiative heat gain. Ecol. Entomol. 2011, 36, 686–699.
- Stabentheiner, A.; Kovac, H. Honeybee economics: Optimisation of foraging in a variable world. Sci. Rep. 2016, 6, 1–7.
- Heinrich, B. Mechanisms of body temperature regulation in honeybees, Apis mellifera. I. Regulation of Head Temperature. J. Exp. Biol. 1980, 85, 61–72.
- Heinrich, B. Mechanisms of body-temperature regulation in honeybees, Apis mellifera. II. Regulation of thoracic temperature at high air temperatures. J. Exp. Biol. 1980, 85, 73–87.
- Attar, I.; Farhat, A. Efficiency evaluation of a solar water heating system applied to the greenhouse climate. Sol. Energy 2015, 119, 212–224.
- Vadiee, A.; Martin, V. Thermal energy storage strategies for effective closed greenhouse design. Appl. Energy 2013, 109, 337–343.
- Ghoulem, M.; El Moueddeb, K.; Nehdi, E.; Boukhanouf, R.; Calautit, J.K. Greenhouse design and cooling technologies for sustainable food cultivation in hot climates: Review of current practice and future status. Biosyst. Eng. 2019, 183, 121–150.
- Zhou, B.; Lin, S.; Jing, S.U.; Xue, F.; Jiang, T. Effects of temperature on the developments of honeybee oosperms and queen pupae. J. Fujian Agric. Univ. 2002, 31, 511–513.
- Zhu, X.; Zhou, B.; Luo, Q.; Luo, F.; Zhong, X.; Shi, M. The effect of temperature on the development of worker honey bee during sealed brood period. New Agric. Technol. 2006, 7, 57–60.
- Tautz, J.; Maier, S.; Groh, C.; Rossler, W.; Brockmann, A. Behavioral performance in adult honey bees is influenced by the temperature experienced during their pupal development. Proc. Natl. Acad. Sci. USA 2003, 100, 7343–7347.
- Groh, C.; Tautz, J.; Rossler, W. Synaptic organization in the adult honey bee brain is influenced by brood-temperature control during pupal development. Proc. Natl. Acad. Sci. USA 2004, 101, 4268–4273.
- Becher, M.A.; Scharpenberg, H.; Moritz, R.F.A. Pupal developmental temperature and behavioral specialization of honeybee workers (Apis mellifera L.). J. Comp. Physiol. A-Neuroethol. Sens. Neural Behav. Physiol. 2009, 195, 673–679.
- Doull, K.M. The effects of different humidities on the hatching of the eggs of honeybees. Apidologie 1976, 7, 61–66.
- Woodrow, W.A. Some Effects of Relative Humidity on the Length of Life and Food Consumption of Honeybees. J. Econ. Entomol. 1935, 28, 565–568.
- Jain, R.; Brockmann, A. Sex-specific molecular specialization and activity rhythm-dependent gene expression in honey bee antennae. J. Exp. Biol. 2020, 223, jeb217406.
- Lacher, V. Elektrophysiologische Untersuchungen an einzelnen Rezeptoren für Geruch, Kohlendioxyd, Luftfeuchtigkeit und Tempratur auf den Antennen der Arbeitsbiene und der Drohne (Apis mellifica L.). Z. Vgl. Physiol. 1964, 48, 587–623.
- Koeniger, N.; Veith, H.J. Glyceryl-1,2-dioleate-3-palmitate, a brood pheromone of the honey bee (Apis mellifera L.). Experientia 1983, 39, 1051–1052.
- Bujok, B.; Kleinhenz, M.; Fuchs, S.; Tautz, J. Hot spots in the bee hive. Naturwissenschaften 2002, 89, 299–301.
- Kleinhenz, M.; Bujok, B.; Fuchs, S.; Tautz, H. Hot bees in empty broodnest cells: Heating from within. J. Exp. Biol. 2003, 206, 4217–4231.
- Kronenberg, F.; Heller, H.C. Colonial thermoregulation in honey bees (Apis mellifera). J. Comp. Physiol. B 1982, 148, 65–76.
- Southwick, E.E. HOGG. Temperature Control in Honey Bee Colonies. BioScience 1987, 37, 395–399.
- Schneider, C.W.; Tautz, J.; Gruenewald, B.; Fuchs, S. RFID Tracking of Sublethal Effects of Two Neonicotinoid Insecticides on the Foraging Behavior of Apis mellifera. PLoS ONE 2012, 7, e30023.
- Henry, M.; Beguin, M.; Requier, F.; Rollin, O.; Odoux, J.-F.; Aupinel, P.; Aptel, J.; Tchamitchian, S.; Decourtye, A. A Common Pesticide Decreases Foraging Success and Survival in Honey Bees. Science 2012, 336, 348–350.
- Campbell, J.M.; Dahn, D.C.; Ryan, D.A.J. Capacitance-based sensor for monitoring bees passing through a tunnel. Meas. Sci. Technol. 2005, 16, 2503–2510.
- Jiang, J.-A.; Wang, C.-H.; Chen, C.-H.; Liao, M.-S.; Su, Y.-L.; Chen, W.-S.; Huang, C.-P.; Yang, E.-C.; Chuang, C.-L. A WSN-based automatic monitoring system for the foraging behavior of honey bees and environmental factors of beehives. Comput. Electron. Agric. 2016, 123, 304–318.
- BeeHero. Available online: https://www.beehero.io/ (accessed on 10 August 2022).
- Huimin, W.; Chuzhou, T.; Ming, L.; Zhongqiu, L.; Zhen, H.; Mingliang, W.; Juying, H.; Haiqing, Z. Effect of airflow speed on pollens distribution for hybrid rice breeding pollination. Trans. Chin. Soc. Agric. Eng. (Trans. CSAE) 2012, 28, 63–69.
- Sedgley, M.; Griffin, A.R. Sexual Reproduction of Tree Crops; Academic Press: Cambridge, MA, USA, 2013.
- Pinillos, V.; Cuevas, J. Artificial pollination in tree crop production. Hortic. Rev. 2008, 34, 239–276.
- Shu, Q.; Liu, Z.; Zhang, J.; Yun, J. Effects of different pollinators on the pollination effect of kiwifruit. J. Zhejiang Agric. Sci. 2015, 56, 1416–1417.
- Jianye, C.; Zhanhong, L.; Yuxia, N. Preparation method and biological effects of pollen suspension liquid forspraying pollination of Actinidia chinensis. J. Fruit Sci. 2014, 31, 1105–1109.
- Cerović, R.; Fotirić Akšić, M.; Meland, M. Success Rate of Individual Pollinizers for the Pear Cultivars “Ingeborg” and “Celina” in a Nordic Climate. Agronomy 2020, 10, 970.
- Santiago, J.P.; Sharkey, T.D. Pollen development at high temperature and role of carbon and nitrogen metabolites. Plant Cell Environ. 2019, 42, 2759–2775.
- Yi, W.; Law, S.E.; McCoy, D.; Wetzstein, H.Y. Stigma development and receptivity in almond (Prunus dulcis). Ann. Bot. 2006, 97, 57–63.
- Mahamad, H.K.; Ali, I. Effects of fungicides on pollen germination peach and nectarine in vitro. Afr. J. Plant Sci. 2011, 5, 643–647.
- Ascari, L.; Guastella, D.; Sigwebela, M.; Engelbrecht, G.; Stubbs, O.; Hills, D.; De Gregorio, T.; Siniscalco, C. Artificial pollination on hazelnut in South Africa: Preliminary data and perspectives. In Proceedings of the IX International Congress on Hazelnut 1226, Samsun, Turkey, 15–18 August 2017; pp. 141–148.
- Parker, A.J.; Tran, J.L.; Ison, J.L.; Bai, J.D.K.; Weis, A.E.; Thomson, J.D. Pollen packing affects the function of pollen on corbiculate bees but not non-corbiculate bees. Arthropod-Plant Interact. 2015, 9, 197–203.
- Wizenberg, S.B.; Weis, A.E.; Campbell, L.G. Comparing methods for controlled capture and quantification of pollen in Cannabis sativa. Appl. Plant Sci. 2020, 8, e11389.
- Vaknin, Y.; Gan-Mor, S.; Bechar, A.; Ronen, B.; Eisikowitch, D. Effects of desiccation and dilution on germinability of almond pollen. J. Hortic. Sci. Biotechnol. 1999, 74, 321–327.
- Buitink, J.; Claessens, M.M.; Hemminga, M.A.; Hoekstra, F.A. Influence of water content and temperature on molecular mobility and intracellular glasses in seeds and pollen. Plant Physiol. 1998, 118, 531–541.
- Payne, W.W. Structure and function in angiosperm pollen wall evolution. Rev. Palaeobot. Palynol. 1981, 35, 39–59.
- Jiayu, H.; Yigang, Z.; Yuanping, C.; Chunqing, W.; Youjin, L. A Strawberry Artificial Pollinator. No. ZL201920082178.3, 24 September 2019.
- Shuo, W.; Jizhan, L.; Jiangshan, W.; Meng, H. A Kind of Pneumatic Strawberry Pollinating Robot in Slightly Raised Greenhouse and Its Realization Method. No. CN109588305A, 9 April 2019.