Polymorphism (rs1805086), c.458A>G, p.Lys(K)153Arg(R), (K153R) of the myostatin gene (MSTN) has been associated with a skeletal muscle phenotype (hypertrophic response in muscles due to strength training). The K153R polymorphism is significant in the development of muscle mass and strength. The rare R variant increases the inhibition of MSTN synthesis, thereby leading to an increase in skeletal muscle mass and muscle strength. The R variant is favorable for sports in which muscle strength and mass are important, such as bodybuilding, powerlifting, weightlifting, arm wrestling, kettlebell lifting, shot put, and bobsleigh. It can be assumed that the strong effect of this allele on the ability to become a successful athlete in weightlifting and speed-power sports is based on the inhibition of MSTN synthesis.
- myostatin
- MSTN
- muscle
- strength
1. History
1.1. MSTN Inhibitors
1.2. Mechanism of Effect of MSTN on Skeletal Muscle Mass and Strength
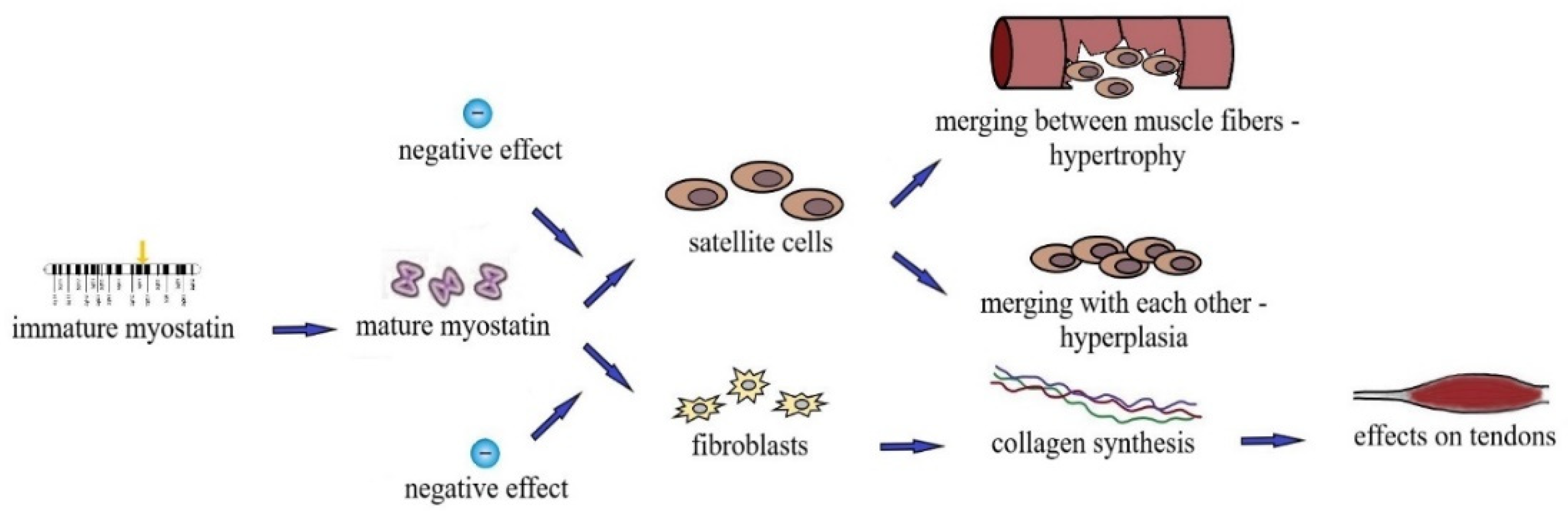
1.3. Effect of MSTN on Tendons and Bones
2. Myostatin Mutations
2.1. MSTN Mutation (rs397515373, c.373 + 5 G>A)
2.2. MSTN A55T Mutation (rs180565, 163 G>A)
2.3. Mutation of MSTN E164K rs35781413 (c.490G>A, p.Glu164Lus)
2.4. Mutation of MSTN K153R (rs1805086, p.Lys153Arg, c.458A>G)
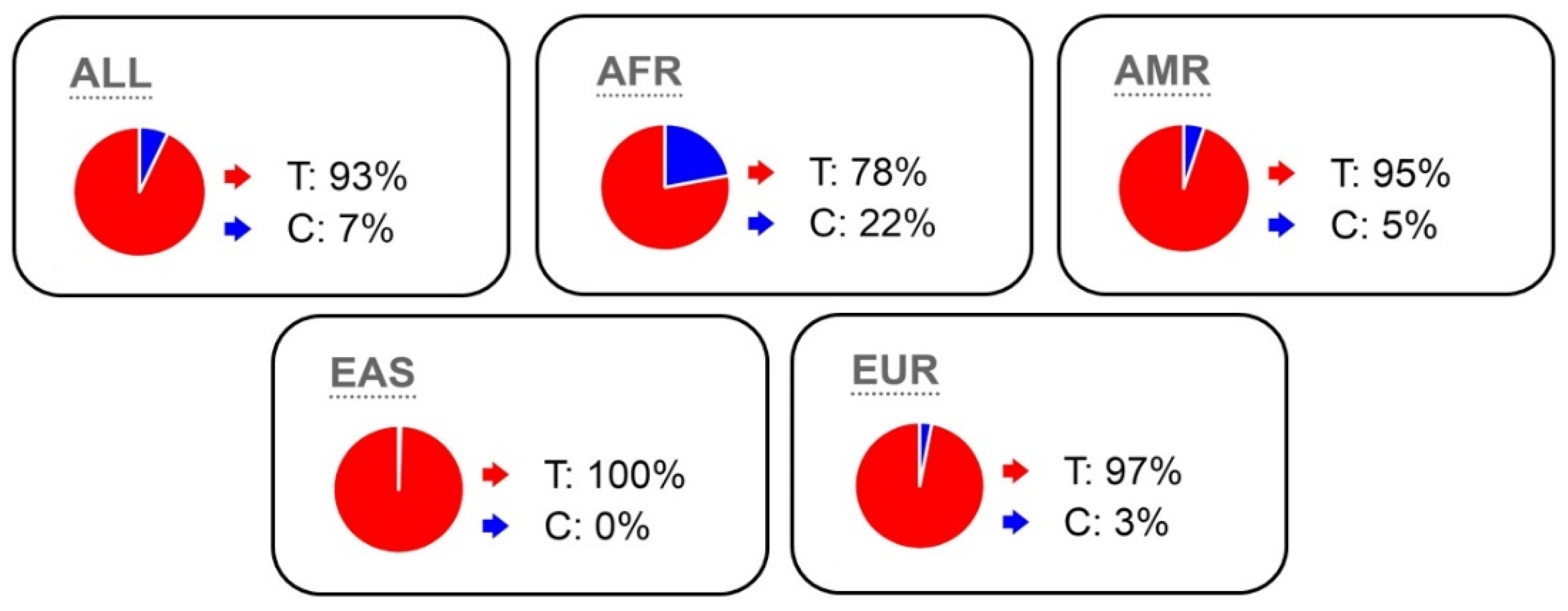
2.5. MSTN K153R (rs1805086) Polymorphism Frequency
This entry is adapted from the peer-reviewed paper 10.3390/genes13112055
References
- Beunen, G.; Thomis, M. Gene powered? Where to go from heritability (H-2) in muscle strength and power? Exerc. Sport Sci. Rev. 2004, 32, 148–154.
- Mangine, G.T.; Hoffman, J.R.; Gonzalez, A.M.; Townsend, J.R.; Wells, A.J.; Jajtner, A.R.; Beyer, K.S.; Boone, C.H.; Miramonti, A.A.; Wang, R.; et al. The effect of training volume and intensity on improvements in muscular strength and size in resistance-trained men. Physiol. Rep. 2015, 3, 17.
- Rodriguez, J.; Vernus, B.; Chelh, I.; Cassar-Malek, I.; Gabillard, J.C.; Sassi, A.H.; Seiliez, I.; Picard, B.; Bonnieu, A. Myostatin and the skeletal muscle atrophy and hypertrophy signaling pathways. Cell. Mol. Life Sci. 2014, 71, 4361–4371.
- Yamada, A.K.; Verlengia, R.; Bueno, C.R. Myostatin: Genetic variants, therapy and gene doping. Braz. J. Pharm. Sci. 2012, 48, 369–377.
- Dalbo, V.J.; Roberts, M.D.; Sunderland, K.L.; Poole, C.N.; Stout, J.R.; Beck, T.W.; Bemben, M.; Kerksick, C.M. Acute Loading and Aging Effects on Myostatin Pathway Biomarkers in Human Skeletal Muscle After Three Sequential Bouts of Resistance Exercise. J. Gerontol. Ser. A-Biol. Sci. Med. Sci. 2011, 66, 855–865.
- Allen, D.L.; Hittel, D.S.; McPherron, A.C. Expression and Function of Myostatin in Obesity, Diabetes, and Exercise Adaptation. Med. Sci. Sport. Exerc. 2011, 43, 1828–1835.
- Zheng, L.-F.; Chen, P.-J.; Xiao, W.-H. Signaling pathways controlling skeletal muscle mass. Acta Physiol. Sin. 2019, 49, 671–679.
- Roth, S.M.; Martel, G.F.; Ferrell, R.E.; Metter, E.J.; Hurley, B.F.; Rogers, M.A. Myostatin gene expression is reduced in humans with heavy resistance strength training: A brief communication. Exp. Biol. Med. 2003, 228, 706–709.
- Shishkin, S.S. Miostatin i nekotorye drugie biohimicheskie faktory, reguliruyushchie rost myshechnyh tkanej u cheloveka i ryada vysshih pozvonochnyh. Uspekhi Biol. Him. 2004, 44, 209–262.
- McPherron, A.C.; Lawler, A.M.; Lee, S.J. Regulation of skeletal muscle mass in mice by a new TGF-β superfamily member. Nature 1997, 387, 83–90.
- Kollias, H.D.; McDermott, J.C. Transforming growth factor-β and myostatin signaling in skeletal muscle. J. Appl. Physiol. 2008, 104, 579–587.
- McFarlane, C.; Hui, G.Z.; Amanda, W.Z.W.; Lau, H.Y.; Lokireddy, S.; Ge, X.J.; Mouly, V.; Butler-Browne, G.; Gluckman, P.D.; Sharma, M.; et al. Human myostatin negatively regulates human myoblast growth and differentiation. Am. J. Physiol.-Cell Physiol. 2011, 301, C195–C203.
- Ferrell, R.E.; Conte, V.; Lawrence, E.C.; Roth, S.M.; Hagberg, J.M.; Hurley, B.F. Frequent sequence variation in the human myostatin (GDF8) gene as a marker for analysis of muscle-related phenotypes. Genomics 1999, 62, 203–207.
- Sergeeva, K.V.; Miroshnikov, A.B.; Smolensky, A.V. Effect of Growth Hormone Administration on the Mass And Strength of Muscles in Healthy Young Adults: A Systematic Review and Meta-Analysis. Hum. Physiol. 2019, 4, 452–460.
- Pan, H.; Ping, X.C.; Zhu, H.J.; Gong, F.Y.; Dong, C.X.; Li, N.S.; Wang, L.J.; Yang, H.B. Association of myostatin gene polymorphisms with obesity in Chinese north Han human subjects. Gene 2012, 494, 237–241.
- Thomis, M.A.; Huygens, W.; Peeters, M.; Vlietinck, R.; Beunen, G.P. Linkage analysis of myostatin-pathway genes in human adiposity: The Leuven Genes for Muscular Strength Project. Med. Sci. Sport. Exerc. 2004, 36, S99.
- Feder, D.; Rugollini, M.; Santomauro, A.; Oliveira, L.P.; Lioi, V.P.; dos Santos, R.; Ferreira, L.G.; Nunes, M.T.; Carvalho, M.H.; Delgado, P.O.; et al. Erythropoietin reduces the expression of myostatin in mdx dystrophic mice. Braz. J. Med. Biol. Res. 2014, 47, 966–971.
- Gentile, M.A.; Nantermet, P.V.; Vogel, R.L.; Phillips, R.; Holder, D.; Hodor, P.; Cheng, C.; Dai, H.Y.; Freedman, L.P.; Ray, W.J. Androgen-mediated improvement of body composition and muscle function involves a novel early transcriptional program including IGF1, mechano growth factor, and induction of β-catenin. J. Mol. Endocrinol. 2010, 44, 55–73.
- Kim, J.S.; Cross, J.M.; Bamman, M.M. Impact of resistance loading on myostatin expression and cell cycle regulation in young and older men and women. Am. J. Physiol.-Endocrinol. Metab. 2005, 288, E1110–E1119.
- Lach-Trifilieff, E.; Minetti, G.C.; Sheppard, K.; Ibebunjo, C.; Feige, J.N.; Hartmann, S.; Brachat, S.; Rivet, H.; Koelbing, C.; Morvan, F.; et al. An Antibody Blocking Activin Type II Receptors Induces Strong Skeletal Muscle Hypertrophy and Protects from Atrophy. Mol. Cell. Biol. 2014, 34, 606–618.
- Jespersen, J.G.; Nedergaard, A.; Andersen, L.L.; Schjerling, P.; Andersen, J.L. Myostatin expression during human muscle hypertrophy and subsequent atrophy: Increased myostatin with detraining. Scand. J. Med. Sci. Sport. 2011, 21, 215–223.
- Lee, S.J.; McPherron, A.C. Regulation of myostatin activity and muscle growth. Proc. Natl. Acad. Sci. USA 2001, 98, 9306–9311.
- Walker, R.G.; Poggioli, T.; Katsimpardi, L.; Buchanan, S.M.; Oh, J.; Wattrus, S.; Heidecker, B.; Fong, Y.W.; Rubin, L.L.; Ganz, P.; et al. Biochemistry and Biology of GDF11 and Myostatin Similarities, Differences, and Questions for Future Investigation. Circ. Res. 2016, 118, 1125–1141.
- Hill, J.J.; Qiu, Y.C.; Hewick, R.M.; Wolfman, N.M. Regulation of myostatin in vivo by growth and differentiation factor-associated serum protein-1: A novel protein with protease inhibitor and follistatin domains. Mol. Endocrinol. 2003, 17, 1144–1154.
- Huang, Z.Q.; Chen, X.L.; Chen, D.W. Myostatin: A novel insight into its role in metabolism, signal pathways, and expression regulation. Cell. Signal. 2011, 23, 1441–1446.
- Drummond, M.J.; Glynn, E.L.; Fry, C.S.; Dhanani, S.; Volpi, E.; Rasmussen, B.B. Essential Amino Acids Increase MicroRNA-499,-208b, and-23a and Downregulate Myostatin and Myocyte Enhancer Factor 2C mRNA Expression in Human Skeletal Muscle. J. Nutr. 2009, 139, 2279–2284.
- Gonzalez-Freire, M.; Rodriguez-Romo, G.; Santiago, C.; Bustamante-Ara, N.; Yvert, T.; Gomez-Gallego, F.; Rexach, J.A.S.; Ruiz, J.R.; Lucia, A. The K153R variant in the myostatin gene and sarcopenia at the end of the human lifespan. Age 2010, 32, 405–409.
- Ben-Zaken, S.; Meckel, Y.; Nemet, D.; Rabinovich, M.; Kassem, E.; Eliakim, A. Frequency of the MSTN Lys(K)-153Arg(R) polymorphism among track & field athletes and swimmers. Growth Horm. IGF Res. 2015, 25, 196–200.
- Fuku, N.; Alis, R.; Yvert, T.; Zempo, H.; Naito, H.; Abe, Y.; Arai, Y.; Murakami, H.; Miyachi, M.; Pareja-Galeano, H.; et al. Muscle-Related Polymorphisms (MSTN rs1805086 and ACTN3 rs1815739) Are Not Associated with Exceptional Longevity in Japanese Centenarians. PLoS ONE 2016, 11, 0166605.
- Joulia-Ekaza, D.; Cabello, G. The myostatin gene: Physiology and pharmacological relevance. Curr. Opin. Pharmacol. 2007, 7, 310–315.
- Kostyunina, D.S.; Ivanova, A.D.; Smirnova, O.V. Myostatin: 20 years later. Hum. Physiol. 2018, 1, 99–114.
- Baczek, J.; Silkiewicz, M.; Wojszel, Z.B. Myostatin as a Biomarker of Muscle Wasting and other Pathologies-State of the Art and Knowledge Gaps. Nutrients 2020, 12, 2401.
- Szlama, G.; Trexler, M.; Buday, L.; Patthy, L. K153R polymorphism in myostatin gene increases the rate of promyostatin activation by furin. FEBS Lett. 2015, 589, 295–301.
- Walsh, F.S.; Celeste, A.J. Myostatin: A modulator of skeletal-muscle stem cells. Biochem. Soc. Trans. 2005, 33, 1513–1517.
- Zhang, Z.L.; He, J.W.; Qin, Y.J.; Hu, Y.Q.; Li, M.; Zhang, H.; Hu, W.W.; Liu, Y.J.; Gu, J.M. Association between myostatin gene polymorphisms and peak BMD variation in Chinese nuclear families. Osteoporos. Int. 2008, 19, 39–47.
- Elkasrawy, M.N.; Hamrick, M.W. Myostatin (GDF-8) as a key factor linking muscle mass and bone structure. J. Musculoskelet. Neuronal Interact. 2010, 10, 56–63.
- Zhu, J.; Li, Y.; Shen, W.; Qiao, C.; Ambrosio, F.; Lavasani, M.; Nozaki, M.; Branca, M.F.; Huard, J. Relationships between transforming growth factor-β 1, myostatin, and decorin—Implications for skeletal muscle fibrosis. J. Biol. Chem. 2007, 282, 25852–25863.
- Guo, W.; Flanagan, J.; Jasuja, R.; Kirkland, J.; Jiang, L.; Bhasin, S. The effects of myostatin on adipogenic differentiation of human bone marrow-derived mesenchymal stem cells are mediated through cross-communication between Smad3 and Wnt/β-catenin signaling pathways. J. Biol. Chem. 2008, 283, 9136–9145.
- Artaza, J.N.; Bhasin, S.; Magee, T.R.; Reisz-Porszasz, S.; Shen, R.Q.; Groome, N.P.; Fareez, M.M.; Gonzalez-Cadavid, N.F. Myostatin inhibits myogenesis and promotes adipogenesis in C3H 10T(1/2) mesenchymal multipotent cells. Endocrinology 2005, 146, 3547–3557.
- Hamrick, M.W.; Arounleut, P.; Kellum, E.; Cain, M.; Immel, D.; Liang, L.F. Recombinant Myostatin (GDF-8) Propeptide Enhances the Repair and Regeneration of Both Muscle and Bone in a Model of Deep Penetrant Musculoskeletal Injury. J. Trauma-Inj. Infect. Crit. Care 2010, 69, 579–583.
- Schuelke, M.; Wagner, K.R.; Stolz, L.E.; Hübner, C.; Riebel, T.; Kömen, W.; Braun, T.; Tobin, J.F.; Lee, S.-J. 2004. Myostatin Mutation Associated with Gross Muscle Hypertrophy in a Child. N. Engl. J. Med. 2004, 350, 2682–2688.
- Corsi, A.M.; Ferrucci, L.; Gozzini, A.; Tanini, A.; Brandi, M.L. Myostatin polymorphisms and age-related sarcopenia in the Italian population. J. Am. Geriatr. Soc. 2002, 50, 1463.
- Li, X.; Wang, S.J.; Tan, S.C.; Chew, P.L.; Liu, L.H.; Wang, L.; Wen, L.; Ma, L.H. The A55T and K153R polymorphisms of MSTN gene are associated with the strength training-induced muscle hypertrophy among Han Chinese men. J. Sport Sci. 2014, 32, 883–891.
- Kostek, M.A.; Angelopoulos, T.J.; Clarkson, P.M.; Gordon, P.M.; Moyna, N.M.; Visich, P.S.; Zoeller, R.F.; Price, T.B.; Seip, R.L.; Thompson, P.D.; et al. Myostatin and Follistatin Polymorphisms Interact with Muscle Phenotypes and Ethnicity. Med. Sci. Sport Exerc. 2009, 41, 1063–1071.
- Kim, J.; Park, K.; Lee, J. Myostatin A55T Genotype is Associated with Strength Recovery Following Exercise-Induced Muscle Damage. Int. J. Environ. Res. Public Health 2020, 17, 4900.
- Juffer, P.; Furrer, R.; Gonzalez-Freire, M.; Santiago, C.; Verde, Z.; Serratosa, L.; Morate, F.J.; Rubio, J.C.; Martin, M.A.; Ruiz, J.R.; et al. Genotype Distributions in Top-level Soccer Players: A Role for ACE? Int. J. Sport Med. 2009, 30, 387–392.
- Aksenov, M.O.; Andryushchenko, L.B. Myostatin gene role in strength building process. Teor. Prakt. Fiz. Kult. 2018, 4, 71–73.
- Santiago, C.; Ruiz, J.R.; Rodriguez-Romo, G.; Fiuza-Luces, C.; Yvert, T.; Gonzalez-Freire, M.; Gomez-Gallego, F.; Moran, M.; Lucia, A. The K153R Polymorphism in the Myostatin Gene and Muscle Power Phenotypes in Young, Non-Athletic Men. PLoS ONE 2011, 6, e16323.
- Filonzi, L.; Franchini, N.; Vaghi, M.; Chiesa, S.; Nonnis Marzano, F. The potential role of myostatin and neurotransmission genes in elite sport performances. J. Biosci. 2015, 40, 531–537.
- Ben-Zaken, S.; Meckel, Y.; Nemet, D.; Eliakim, A. The combined frequency of IGF and myostatin polymorphism among track & field athletes and swimmers. Growth Horm. IGF Res. 2017, 32, 29–32.
- Khanal, P.; He, L.X.; Herbert, A.J.; Stebbings, G.K.; Onambele-Pearson, G.L.; Degens, H.; Morse, C.I.; Thomis, M.; Williams, A.G. The Association of Multiple Gene Variants with Ageing Skeletal Muscle Phenotypes in Elderly Women. Genes 2020, 11, 1459.
- Grealy, R.; Herruer, J.; Smith, C.L.E.; Hiller, D.; Haseler, L.J.; Griffiths, L.R. Evaluation of a 7-Gene Genetic Profile for Athletic Endurance Phenotype in Ironman Championship Triathletes. PLoS ONE 2015, 10, e0145171.