2. Oxidative Stress
Oxidative stress occurs when RONS overwhelm cellular defenses, causing damage to proteins, membranes, and deoxyribonucleic acid (DNA) [
35]. It is the result of a disturbance in the balance between RONS and antioxidants in favor of RONS [
36]. Under physiological conditions, endogenous RONS are generated by enzymatic systems, including nicotinamide adenine dinucleotide phosphate oxidase (NADPH oxidase) and nitric oxide synthase (NOS), as a by-product of mitochondrial electron transport chain reactions (
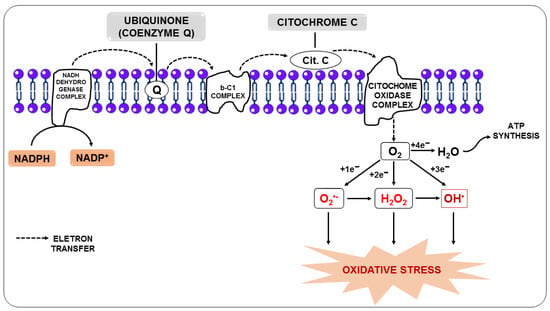
Figure 1) or by metal-catalyzed oxidation [
37,
38].
Figure 1. Production of reactive oxygen species from the transfer of electrons from the electron transport chain.
In this regard, the free radical superoxide (O
2•−), resulting from the monoelectronic reduction of oxygen, is considered the main precursor of other RONS since, after its formation, it can react with other molecules giving rise to other free radicals, such as hydroxyl (OH
•), alkoxyl (RO
-), and peroxyl (ROO
-), in addition to other molecules that do not meet the definition of free radicals, but take part of oxidative reactions in a meaningful way, such as H
2O
2. Nitric oxide (NO) is among the molecules that can react with O
2•−, and the reaction between them generates the free radical peroxynitrite (ONOO
-). Additionally, O
2•− can be unmuted to form H
2O
2, and it can be broken down through Fenton or Haber-Weiss reactions, leading to the generation of OH
• [
39,
40].
These RONS-generating chain reactions are initially controlled by antioxidant defense systems that act quickly, neutralizing any molecule that can potentially develop into a RONS or any free radical with the ability to induce the production of other pro-oxidants [
41]. Three enzymes are critical in this process, including superoxide dismutase (SOD), catalase (CAT), and glutathione peroxidase (GSH-Px). These enzymes, respectively, unmute O
2•− and break down H
2O
2 or hydroperoxides (ROOH) into harmless molecules such as H
2O, alcohol, and oxygen (O
2) [
42]. The class of endogenous antioxidants also includes glutathione reductase, and reduced glutathione (GSH), in addition to small molecules such as coenzyme Q and uric acid (UA), among others [
43]. Since they can be synthesized by the body in response to oxidative aggression, the researchers nominate endogenous antioxidants as
mobilizable antioxidant molecules.
However, in diseases in which oxidative stress is a pathogenic mediator, including cancer and malaria, mobilizable antioxidants are not sufficient to maintain cell homeostasis due to the decreased synthesis of antioxidant enzymes and increased use of these antioxidants, among other factors [
44,
45,
46].
In these cases, supplementation with dietary antioxidants is essential to maintain optimal cell function. Vitamins, including vitamins E and C, phenolic substances, such as flavonoids, resveratrol, and carotenoids, including β-carotene and lycopene, and drugs, such as
N-acetylcysteine (NAC), among others, belong to this category [
47,
48]. Dietary antioxidants neutralize or eliminate RONS by binding or donating electrons to pro-oxidants, and in the process, they become free radicals but with less harmful effects. These “new radicals” are more easily neutralized and rendered completely harmless by other antioxidants in this group [
49]. Thus, this class of antioxidant molecules can also be referred to as
consumable antioxidants, as they are consumed in the face of oxidative aggression. Thus, consumable and mobilizable antioxidants act synergistically to fight the excessive increase in RONS, which can be a primary cause or a secondary complication of various diseases [
50,
51], as in malaria [
52,
53].
3. Oxidative Stress in Malaria
In malaria, oxidative stress is caused by four main mechanisms: a host defense against
Plasmodium infection; ischemia-reperfusion syndrome; direct production of oxidative species by the parasite; and the metabolism of antimalarial drugs [
54].
3.1. Oxidative Stress as a Host Defense Mechanism
RONS are essential for several physiological functions of the body, including cell survival, growth, proliferation, and differentiation, as well as the immune response [
55,
56].
As for the immune response, RONS are important for phagocytes, including neutrophils and monocytes/macrophages, which are highly activated during malaria, helping these cells phagocytize and destruct parasites [
57,
58].
In this sense, the body’s defense system responds to infection by primarily recruiting neutrophils [
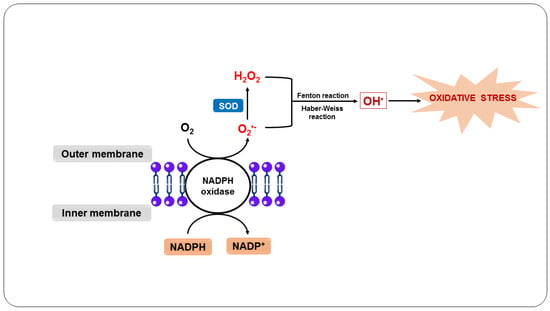
59]. When neutrophils engulf the parasites, they induce a respiratory burst (
Figure 2), in which O
2 enzymatically reacts with NADPH oxidase present in the plasma and the phagosomal membrane of neutrophils, forming O
2•− [
4]. O
2•− and its derivatives H
2O
2 and OH
•, when released by activated neutrophils in the phagosome, are essential to kill ingested pathogens [
60].
Figure 2. Oxidative stress as a host defense mechanism in response to infection by Plasmodium sp.
In addition, activated neutrophils produce cytokines, such as GM-CSF and M-CSF, and chemokines, including MIP-1α and MIP-1β, which attract these cells and are essential for monocyte mobilization [
61]. These leukocytes engulf and kill the parasites through the oxidative action of O
2•−, which is generated in the same way as in neutrophils, as well as by the action of NO, which is produced by the macrophage from the reaction of NOS with L-arginine [
62,
63]. Additionally, the NO and O
2•− generated react to form other RONS, such as ONOO
-, intensifying the cytotoxicity directed against the parasites [
64].
Furthermore, neutrophil and macrophage myeloperoxidase is activated and uses H
2O
2 as a substrate to produce hypochlorous acid, a highly bactericidal compound [
65,
66]. On the other hand, phagocytosis and the consequent action of RONS, including O
2•− and NO, as well as other toxic products, can exacerbate the condition due to rupture of the parasitized erythrocytes, during which normal uninfected erythrocytes can also be destroyed, stimulating cytoadherence and, consequently, potentially blocking blood flow, causing ischemia and anemia [
67,
68].
3.2. Oxidative Stress Due to Ischemia-Reperfusion Syndrome
In individuals with malaria, severe anemia induces microvascular dysfunction, leading to recurrent episodes of initial restriction of blood supply to organs, which can lead to ischemia and nutrient and oxygen deprivation, followed by subsequent restoration of concurrent perfusion and reoxygenation [
54,
69]. This process is called ischemia-reperfusion syndrome and can occur in malaria due to the sequestration of parasitized erythrocytes, as a result of the destruction of erythrocytes caused by the parasites and RONS during the paroxysm of malaria, and due to cytoadherence of erythrocytes to blood vessels [
70].
Furthermore, this syndrome can trigger anaerobic metabolism, the production of lactic acid, and the consequent depletion of adenosine triphosphate (ATP). As ATP availability is reduced, ATP-dependent ion channels begin to fail. At the same time, calcium overload and excessive RONS production open the mitochondrial permeability transition pore, further reducing ATP levels [
71,
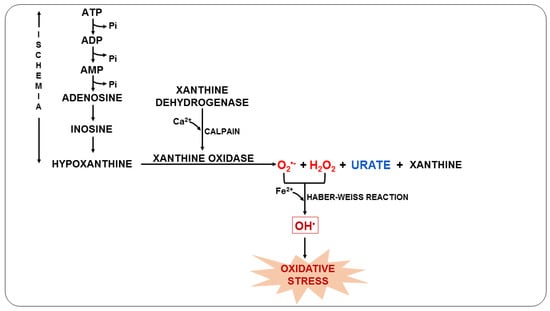
72]. During the ischemic process, the degradation of ATP causes the accumulation of xanthine oxidase (XO) and hypoxanthine due to the lack of oxygen. When the blood supply is resumed, XO acts on hypoxanthine resulting in the production of O
2•− (
Figure 3), which can later be converted into OH
• in the presence of transition metals and, consequently, trigger oxidative stress [
73,
74].
Figure 3. Oxidative stress due to ischemia-reperfusion syndrome during malaria.
During this process, UA is also formed, which is a weak organic acid present mainly as monosodium urate at physiological pH [
75]. UA can be found in the host organism during malarial infection and can act by eliminating RONS and chelating transition metal ions or even by reducing NOS expression, impairing NO release [
76,
77]. Previous studies have shown that plasma UA levels in
P. falciparum-infected children increase during acute episodes and with disease severity, suggesting that UA is an important mediator in the pathophysiology of malaria [
78,
79].
In the ischemia-reperfusion syndrome, RONS can be produced during ischemia but is massively increased during reperfusion, amplifying and propagating oxidative damage and destroying the integrity of proteins, membranes, and microvascular endothelium [
80].
3.3. Oxidative Stress Due to the Metabolism of the Parasite
Another important oxidative mechanism in malaria is mainly triggered by the metabolism of the parasite, as well as by the potentially oxidative by-products generated and released from red blood cells destroyed by the action of the parasite [
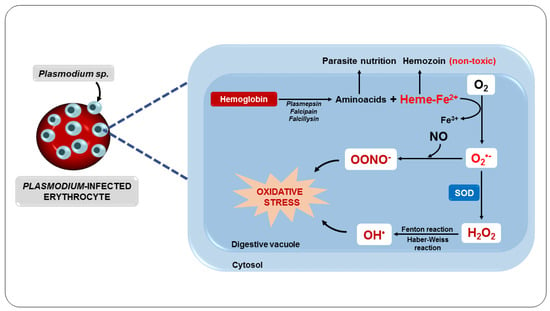
81]. Inside the erythrocyte, the parasite digests hemoglobin in its acidic digestive vacuole, forming essential amino acids for parasite development and proliferation [
82]. However, in this process, ferroprotoporphyrin IX or heme complex (FPIX) is released, which is toxic to the parasite. On the other hand, this complex can still be detoxified within the parasite by polymerization [
83].
Although the parasite manages to polymerize FPIX, resulting in a nontoxic derivative, hemozoin, also known as a malarial pigment, a significant amount escapes polymerization [
84]. Thus, the ferrous iron (Fe
2+) from FPIX is oxidized to the ferric state (Fe
3+), with the consequent production of superoxide, which dismutates to H
2O
2 (
Figure 4). This oxidative reaction chain leads to the production of OH
• from reactions involving H
2O
2 and Fe
3+, such as the Fenton and Haber–Weiss reactions [
85].
Figure 4. Oxidative stress as a consequence of parasite metabolism.
These free radicals can cause damage to the parasite’s digestive vacuole membrane, eventually killing it [
81]. However, the rapid development and proliferation of the parasite, associated with the RONS generated and released inside the erythrocytes, cause structural damage to the erythrocytes [
86]. This results in increased membrane permeability for ions, increased cell volume, oxidation of sulfhydryl groups, and reduced deformability, contributing to the loss of erythrocyte function and cell lysis [
67,
87].
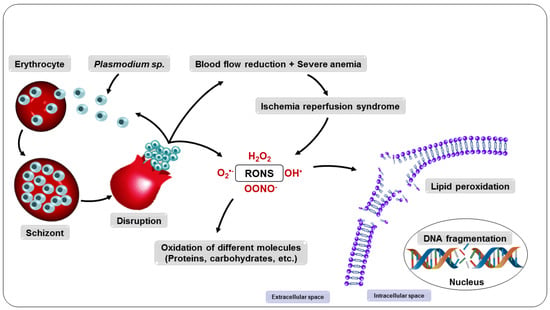
Consequently, all intra-erythrocyte content, including RONS, will be released to the extracellular environment, resulting in damage to several biomolecules, such as lipids, proteins, and DNA, as well as enzyme inactivation, apoptosis induction, modification of surface adhesion molecule expression of leukocytes and endothelial cells, and alteration in the bioavailability of NO, compromising homeostasis and, ultimately, its survival [
88,
89]. These changes expose the host organism to a highly oxidative environment (
Figure 5), implying the development of systemic complications such as reduced blood flow and severe anemia and also facilitating the entry of parasites into tissues such as the lung and brain, which can lead to organ failure [
83,
90,
91,
92].
Figure 5. Consequences of the multiplication of parasites in the erythrocyte.
3.4. Oxidative Stress as a Consequence of the Metabolization of Antimalarial Drugs
The drug treatment of malaria is specially designed to interrupt parasite proliferation, responsible for the pathogenesis and clinical manifestations of the infection, to destroy the latent forms of the parasite (hypnozoites) to prevent late relapses, and to prevent the transmission of the parasite, through the use of drugs that prevent the development of sexual forms of the parasites [
93,
94].
In this context, one of the main targets of antimalarial drugs is the intracellular pathway of heme metabolism, which is implicated in the production of RONS and the consequent death of the parasite [
95,
96]. Therefore, chloroquine, a quinoline blood schizonticidal drug used to treat severe and uncomplicated cases of malaria, can act by preventing FPIX polymerization, causing the accumulation of FPIX in the parasite’s digestive vacuole and consequent lethal oxidative stress in the parasite [
97,
98,
99]. However, there are increasing reports of
P. falciparum resistance to quinoline antimalarials, highlighting the importance of the
P. falciparum chloroquine resistance transporter, a member of the drug/metabolite transporter superfamily located in the parasite’s digestive vacuole, as the main responsible for chloroquine resistance [
100,
101,
102].
Other studies indicate that, in addition to showing chemical similarity with chloroquine and a similar mechanism of action, other quinolines, such as quinine, amodiaquine, lumefantrine, and mefloquine are effective against many strains of parasites resistant to chloroquine [
103,
104,
105]. In addition, some of these drugs are widely used in combination therapies with artemisinin derivatives, including artemether plus lumefantrine and artesunate plus amodiaquine, and provide synergistic antimalarial activity along with preventing the development of antimalarial drug resistance [
106,
107,
108].
The site of action of artemisinin and its derivatives dihydroartemisinin, artemether, arteether, and artesunate is believed to be the parasite’s digestive vacuole, where these drugs can interfere with the FPIX complex, giving rise to RONS, leading to damage to nearby proteins, and still interacting with the mitochondrial electron transport chain of the parasite, enhancing RONS production, impairing mitochondrial functions, and killing the parasite [
109,
110,
111].
Artemisinins act quickly and are very potent against blood-stage parasites. They are active against the sex stages of the parasite, which is important for blocking transmission [
112,
113]. However, due to their short half-life, these drugs are used in conjunction with other long-acting drugs that remain in the body for longer to fight potential remaining parasites [
114,
115].
Accordingly, studies show that primaquine increases the effect of combination therapy with artemisinin derivatives in eliminating malaria and reduces the risk of artemisinin-resistant infections [
116].
Furthermore, only primaquine is recognized for completely eliminating
P. vivax and
P. ovale that form hypnozoites—the latent form of the parasite that remains in the liver and is responsible for disease relapse in individuals infected by these parasites—refractory to most drugs and for providing a radical cure [
93,
117]. Primaquine, an 8-aminoquinoline, can act directly on erythrocytes leading to massive production of RONS and consequent lipid peroxidation of the cytoskeleton and membrane, as well as hemolysis [
118]. However, the use of primaquine in individuals with glucose-6-phosphate dehydrogenase deficiency can result in clinical manifestations of hemolysis, such as severe anemia, fatigue, jaundice, and acute renal failure, thereby limiting its use [
119,
120].
In this scenario, as a product of the normal host’s metabolism or from the metabolism of the parasite, or as an effect of pharmacological treatment, intensely produced RONS cause damage to lipids, proteins, and DNA, leading to oxidative stress that impairs the normal functioning of the infected organism [
98]. Therefore, the search for adjuvant therapies that can improve the clinical outcomes of malaria continues because, despite their benefits, treatments eventually cause oxidative damage, which limits their use [
121].
3.5. Nitric Oxide in Malaria
Scientific evidence demonstrates that a specific RONS is particularly involved in the pathophysiology of this disease: NO [
122,
123]. It has been suggested that the low bioavailability of NO promotes oxidative stress in tissues such as the brain and lungs [
124]. On the other hand, it has been shown that NO at high concentrations can kill
Plasmodium [
125,
126]. NO is an important mediator of biological processes such as vascular homeostasis, neurotransmission, immunity, and inflammation [
127,
128,
129]. Furthermore, it is a free radical produced by three different nitric oxide synthase enzymes, neuronal NOS (nNOS or NOS1), endothelial NOS (eNOS or NOS3), which are constitutively expressed, and the inducible NOS (iNOS or NOS2), which is induced by inflammatory stimuli [
130,
131,
132,
133]. NO is very reactive and has a very short half-life. For this reason, nitrite and nitrate measurements, which are the final metabolites of NO, have been used to measure the concentration of NO indirectly [
134,
135].
Experimental evidence indicates that NO plays an important role in the defense against plasmodia in vitro and in vivo [
136,
137]. In this context, studies have shown that circulating levels of nitrite and nitrate were higher in anopheline mosquitoes—a natural vector of malaria in humans—infected with
Plasmodium and that increased NO concentrations at the beginning of the sporozoite stage induced the formation of toxic metabolites, limiting parasite development [
138].
In children and adults with malaria, elevated plasma levels of nitrites and nitrates have been associated with more rapid parasite clearance [
139]. Indeed, previous studies have shown that children infected with
P. falciparum had elevated levels of NO and iNOS activity, suggesting the protective role of NO in children with malaria [
140]. Protection against severe malaria in this population of children appears to be associated, at least in part, with a polymorphism in the iNOS gene, which produces high levels of NO during an inflammatory event [
141]. These studies suggest that NO production during malaria depends on the severity of the disease and the degree of patient immunity [
142].
In an animal model of experimental cerebral malaria (ECM), Serghides et al. [
143] demonstrated that pretreatment with inhaled NO reduced the accumulation of parasitized erythrocytes in the brain, decreased endothelial cell expression, and preserved vascular integrity. From these results, the authors suggested that prophylaxis with NO inhalation can reduce systemic inflammation and endothelial activation during ECM. In a similar model, Ong et al. [
144] showed that cerebrovascular dysfunction is characterized by vascular constriction, occlusion, and cell damage, resulting in impaired perfusion and reduced cerebral blood flow and oxygenation, and was associated with low NO bioavailability.
Given the critical importance of NO-derived and -non-derived oxidative stress in the underlying pathophysiological mechanisms of the disease, studies have shown that natural or synthetic exogenous antioxidants, including vitamin A, E, zinc, selenium, NAC, curcumin,
Agaricus sylvaticus mushroom, and carotenoids, can benefit the treatment of malaria [
145,
146,
147,
148]. Several studies have indicated an association between the use of carotenoids and a decrease in oxidative changes, suggesting that the antioxidant properties of these compounds are an important factor against malaria-induced oxidative stress [
149,
150]. The recent interest in carotenoids has focused on the role of lycopene in human health [
151,
152].
4. Lycopene
Lycopene is a natural constituent synthesized by plants and microorganisms [
153]. It is a red pigment found in some fruits and vegetables, such as guava, watermelon, papaya, pitanga
(Eugenia uniflora—Myrtaceae), tomatoes, and their derivatives [
154,
155,
156,
157] and can be extracted from these vegetables by chemical reactions using organic solvents, such as ethanol and ethyl acetate and/or using a supercritical fluid such as supercritical carbon dioxide, or by heat treatment at different temperatures ranging from 60 to 140 °C [
158,
159,
160,
161,
162,
163]. It is widely used as a supplement in functional foods, nutraceuticals, and pharmaceuticals, as well as an additive in cosmetics [
164,
165].
Lycopene is an intermediate product of the β-carotene biosynthetic pathway that does not have provitamin A activity, as it does not have the β-ionone ring in its structure, which is responsible for this characteristic [
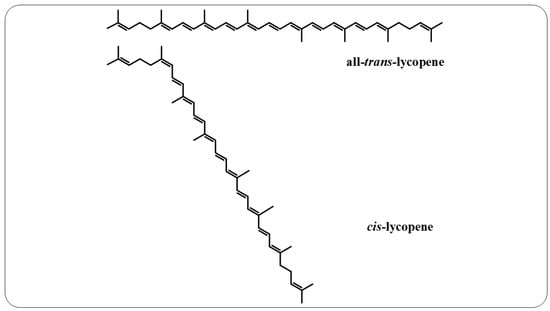
166]. This compound is a noncyclic, fat-soluble hydrocarbon that contains 11 conjugated double bonds and 2 unconjugated double bonds, thereby offering it greater reactivity. This polyene can also exist in all-
trans and
cis-lycopene isomeric forms (
Figure 6). Conversion from all-
trans- to
cis-lycopene forms can occur by geometric isomerism induced by light, thermal energy, or chemical reactions [
167,
168].
Figure 6. All-trans-lycopene and cis-lycopene structures.
5. Antioxidant Effects of Lycopene
Among the carotenoids, lycopene is the most effective antioxidant against RONS and may contribute to preventing or reducing oxidative damage to cells and tissues in vivo and in vitro [
203]. Evidence supports the role of lycopene as a potent antioxidant, capable of scavenging singlet oxygen (
1O
2) and other free radicals, such as ROO
-, with a potential twice as high as β-carotene, and ten times as efficient as α -tocopherol, although lycopene circulates at much lower concentrations than vitamin E [
204,
205]. During the elimination of
1O
2, energy is transferred from this radical to the lycopene molecule and, as it has an open chain with 11 conjugated double bonds in its structure, this favors stabilization of the unpaired electron of the radical by resonance [
206,
207]. Additionally, it was observed that lycopene effectively eliminates other RONS, such as OH
•, O
2•−, and ONOO
- [
208].
Furthermore, the lipophilic characteristic of lycopene favors its interaction with the lipid bilayer of the cell membrane, thereby preventing the breakdown of fatty acids and the oxidation of lipids, proteins, and DNA [
180]. In this sense, Suwannalert et al. [
209], investigating serum levels of lycopene and malondialdehyde (MDA) in elderly susceptible to oxidative stress, demonstrated that lycopene levels were inversely related to MDA levels. Additionally, Yonar and Sakin [
206] demonstrated that lycopene treatment prevented deltamethrin-induced oxidative stress by decreasing MDA levels in fish (
Cyprinus carpio) and significantly increasing SOD, CAT, and GSH-Px activities and the level of GSH. Similar results were found by Kujawska et al. [
210], who reported that treatment with tomato extract enriched with lycopene was able to suppress the oxidative stress induced by
N-nitrosodiethylamine in rats and increase the enzymatic antioxidant activity in these animals.
6. Effects of Lycopene on Malaria
The use of antioxidant compounds in the treatment of tropical diseases has increased, including Chagas disease, dengue, and malaria, as several studies have suggested the involvement of oxidative stress in the pathogenesis and progression of these diseases [
54,
258,
259]. In this context, studies show that the discovery of new antimalarial drugs is necessary, and natural antioxidant products are important sources for obtaining new antimalarial compounds or even as adjuvant therapy, enhancing the activity of antimalarial drugs [
260,
261,
262,
263,
264].
A study by Metzger et al. [
34] demonstrated that natural products can be used in malaria chemotherapy. According to this study, increased plasma lycopene concentration was associated with faster clearance of parasites in children. In a related study, Caulfield et al. [
265] demonstrated that the nutritional deficiency of the host is associated with the morbidity and mortality of children with severe malaria. In this sense, previous studies suggest that changes in plasma concentrations of micronutrients, including vitamins A and C, retinol, β-carotene, α-carotene, β-cryptoxanthin, lutein, and lycopene, occur due to increased use of these antioxidants in patients with malaria, suggesting that there may be a need for vitamin supplementation in patients with malaria [
266]. In corroborating this suggestion, the nutritional deficit seems to be associated with a redirection of these antioxidants to the liver to aid in the synthesis of acute-phase proteins in other organs, repair tissue damage caused by the infectious organism, and aid in the host’s oxidative defense mechanisms [
33].
In fact, Sondo et al. [
32] had already reported that periodic supplementation of high doses of vitamin A and zinc could reduce the morbidity caused by malaria. In this sense, Agarwal et al. [
147] investigated the effect of lycopene on the growth of
P. falciparum in vitro, monitoring the progression at different stages. These authors showed that lycopene treatment induced an increased production of RONS in the cytoplasm of the parasite, which caused the parasite to lose its mitochondrial membrane potential and cytotoxicity, resulting in merozoites not being released from the erythrocytes of the host, suggesting that the inclusion of lycopene in the diet may be useful in changing the clinical outcomes of malaria.
Preliminary results from researchers' group demonstrated that lycopene supplementation in mice (BALB/c; 3.11 mg/kg) infected with the
P. berghei strain showed a delay in the induction and a decreased progression of parasitemia. Also, the animals supplemented with lycopene showed a higher rate of survival compared to the positive control [
267], suggesting lycopene prophylactic and antiparasitic activity, which may be due to the cytotoxic effect of lycopene against the parasite [
147], suggesting an important role of lycopene supplementation in preventing malaria [
268].