Your browser does not fully support modern features. Please upgrade for a smoother experience.
Please note this is an old version of this entry, which may differ significantly from the current revision.
Subjects:
Chemistry, Medicinal
Pyrroloiminoquinones are a group of cytotoxic alkaloids most commonly isolated from marine sponges. Structurally, they are based on a tricyclic pyrrolo[4,3,2-de]quinoline core and encompass marine natural products such as makaluvamines, tsitsikammamines and discorhabdins.
- makaluvamine
- damirone
- discorhabdin
- batzelline
- tsitsikammamine
1. Introduction
Pyrroloiminoquinones are a large and diverse group of natural products that have been isolated predominantly from marine sponges [1,2,3,4]. They are considered to be potential drug leads due to their significant inhibition of cell proliferation in various cancer cell lines, including promising in vivo activity against several tumor types [5,6,7,8,9] and inhibition of Plasmodium berghei parasitemia [10] in mouse models. In addition, pyrroloiminoquinones have been shown to exhibit antiviral [11,12], antifungal [13,14,15] and antibacterial [5,12,15,16,17] as well as neuromodulatory [18,19] and antioxidant [20] activities. The mechanisms of bioactivity for these compounds are not yet completely understood with members of this compound class appearing to exert their activity through a number of different modes of action which, for anticancer cell activity, include direct DNA damage [7,8] and inhibition of key cell regulatory enzymes [21].
Pyrroloiminoquinone molecular structures are characterized by a condensed tricyclic pyrrolo[4,3,2-de]quinoline core that is also considered the principal pharmacophore of this compound class responsible for their antiproliferative and cytotoxic effects [7,22]. Most compounds of this class can be assigned to one of three major classes exhibiting distinct core structures, namely makaluvamines, bispyrroloiminoquinones and discorhabdins (Figure 1).
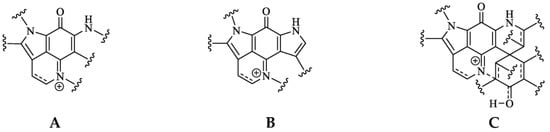
Figure 1. Pyrroloiminoquinone structural scaffolds (A)—Makaluvamines, (B)—Bispyrroloiminoquinones, (C)—Discorhabdins.
Pyrroloiminoquinones have been mostly isolated from marine sponges of the order Poecilosclerida, with Latrunculiidae species from temperate and cold-water environments such as New Zealand, South Africa, the Arctic, and Antarctic as well as warm-water Acarnidae species from the Indo-Pacific proving particularly productive sources [2,3,4]. Nevertheless, members of this class of alkaloids have been reported from ascidians [23,24] and simple representatives have also been isolated from cultured myxomycetes [25,26]. Moreover, closely related alkaloids have been reported in hydroids [27,28], terrestrial fungi [29,30,31,32] and marine actinobacteria [33,34]. This wide geographical and phylogenetic distribution of pyrroloiminoquinone producers, as well as the production of related compounds by bacteria raises the question of microbial involvement in their biosynthesis within marine invertebrates.
2. Structures and Host Distribution of Natural Pyrroloiminoquinones and Related Compounds
2.1. Makaluvamines, Bispyrroloiminoquinones and Discorhabdins
Structurally, the simplest pyrroloiminoquinones are represented by the makaluvamines consisting of the characteristic pyrrolo[4,3,2-de]quinoline core and variable substituents (Figure 2). These include N-methylation of the pyrrole or imine nitrogen, halogenation at C-6, Δ3,4-desaturation and alkylation of N-7 with phenylethyl based side chains. Notable exceptions are makaluvamine O and makaluvamine W and the broad use of the term ‘makaluvamine’ in this review excludes these two structures. Makaluvamines were first reported in 1993 in the sponge Zyzzya fuliginosa collected near the Makaluva Islands, Fiji [7] and since then, have most routinely been isolated from Pacific and Indo-Pacific warm-water sponges of the genus Zyzzya (family Acarnidae) [8,10,11,20,38,39,40,41,42,43,44,45,46,47]. Makaluvamines have also been isolated from latrunculid sponge species collected off South Africa [22,48,49], the Korean peninsula [16], New Zealand [50] and Australia [21]. Interestingly, simple makaluvamines bearing either no substituents or only exhibiting N-methylation have been purified from cultured myxomycetes, Didymium iridis and Didymium bahiense, isolated from Japanese forest litter samples [25,26]. Makaluvamines are thought to be the biosynthetic precursors to more complex pyrroloiminoquinones and the sulfur-containing makaluvamine F may represent a precursor to sulfur-containing discorhabdins [50].
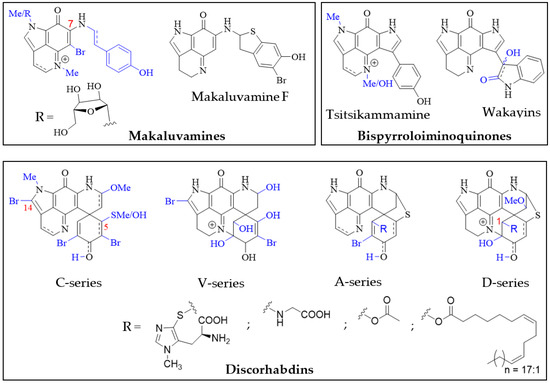
Figure 2. Main pyrroloiminoquinone classes. Variable substituents encompassing all known compound class members are shown in blue.
Bispyrroloiminoquinones are relatively rare pyrroloiminoquinones containing a characteristic pyrrolo[4,3,2-de]pyrrolo[2,3-h]quinoline core and, depending on the nature of the side-chain, they can be classed as either tsitsikammamines or wakayins (Figure 2). Tsitsikammamines have been reported in the South African marine sponges Tsitsikamma favus and Tsitsikamma nguni [15,22,49,51,52], an Australian Zyzzya sp. [45], Tongan Strongylodesma tongaensis [53] as well as Antarctic Latrunculia biformis [54]. Wakayins (wakayin and 16-hydroxy-17-oxyindolewakayin) on the other hand, have only ever been found in ascidians of the genus Clavelina collected in Micronesia (Wakaya Islands) [23] and Thailand [24].
Pyrroloiminoquinones comprising a pyrido[2,3-h]pyrrolo[4,3,2-de]quinoline core with an additional spiro-fused cyclohexanone/-ol or cyclohexadienone/-ol moiety are known as discorhabdins (Figure 2), some of which are historically also referred to as epinardins or prianosins [55,56,57]. Members of this structurally complex and diverse class of pyrroloiminoquinones are known for particularly potent bioactivities and have attracted significant interest from natural product, medicinal and synthetic chemists alike [5,6,7,9,12,14,15,16,18,21,22,35,36,37,54,58,59,60,61,62,63,64,65,66,67,68,69,70,71,72,73,74,75]. In contrast to makaluvamines and bispyrroloiminoquinones, discorhabdins have been exclusively found in demosponges, mostly belonging to the family Latrunculiidae. While they all share a characteristic spiro-arrangement flanking the pyrroloiminoquinone core, most known discorhabdins can be divided into distinct structural sub-classes: First, the often multi-brominated, pentacyclic discorhabdins of the C-series and analogous hexacyclic discorhabdins of the V-series exhibiting N(18)-C(2) ring closure and, second, the hexacyclic discorhabdins of the A-series with a bridging sulfur atom connecting C-5 and C-8 and analogous heptacyclic discorhabdins of the D-series displaying N(18)-C(2) ring closure. Some sponges, such as those of genus Tsitsikamma [15,22,49,52], produce only C- and V-series discorhabdins, while the presence of A- and D-series discorhabdins is often accompanied by discorhabdins of the former classes. C-14 bromination also appears to be particular to Tsitsikamma sponges [15,22,49,52] and may represent a chemotaxonomic distinction, while C-5 thiomethylation has only been observed in isolates from a single Caribbean deep-water Strongylodesma purpureus sponge [76] (reassigned from Batzella [77]). A- and D-series discorhabdins often exhibit substitution at C-1 with substituents such as ovothiol, glycine and alkyl esters [22,66,68,72], possibly as a result of increased electrophilic reactivity of the parent compounds [70]. Furthermore, A- and D-series discorhabdins are chiral and enantiomers of opposite parity have been isolated from the same Latrunculia species collected in different locations [65]. To date, all evidence available for comparison suggests that enantiomeric parity does not significantly affect biological activity [65,72].
2.2. Unusual Pyrroloiminoquinones and Related Pyrroloquinolines from Marine Sponges
In addition to the monomeric discorhabdins discussed above, several dimeric and trimeric discorhabdins have been isolated from sponges of the genus Latrunculia (Figure 3). These comprise W-series discorhabdins characterized by a disulfide bridge linking two discorhabdin monomers [64,66], thioether-linked discorhabdin dimers [70,74], a C-N-linked discorhabdin C dimer [73], a C-N-linked discorhabdin B dimer as well as a discorhabdin trimer [75]. The saturated discorhabdin W dimer and its monomers have been shown to be interconvertible through reductive cleavage and subsequent UV-irradiation [64], whereas thioether-linked discorhabdin dimers were first discovered as a major degradation product of monomeric discorhabdin stored at −20 °C for a fortnight [70]. Such non-enzymatic dimerization together with the observations that discorhabdins have been shown to be prone to nucleophilic attack at C-1 [70,73] suggest that at least sulfur-bridged discorhabdin dimers may be generated non-enzymatically in situ [73]. Furthermore, LC-MS/MS-driven molecular networking has provided evidence for numerous discorhabdin di- and trimers, some even incorporating makaluvamines, alongside monomeric A- and D-series discorhabdins in extracts of subantarctic Latrunculia apicalis and South African Cyclacanthia bellae [52].
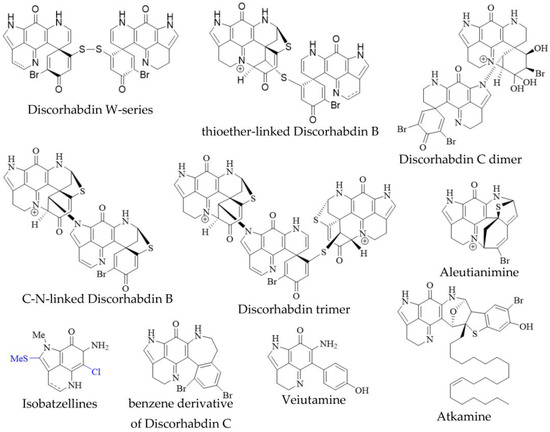
Figure 3. Unusual pyrroloiminoquinones from marine sponges. Variable substituents are shown in blue.
Alike to oligomeric discorhabdins exemplifying a special case of discorhabdin structures, isobatzellines are makaluvamine-like structures that are distinguished by thiomethylation at C-2 or chlorination at C-6 and can thus be regarded as a subgroup of makaluvamines (Figure 3). Isobatzellines exhibiting thiomethylation have been exclusively reported in a Grand Bahaman Strongylodesma nigra specimen [13] (reassigned from Batzella [77]), while those only containing chlorine substituents have also been purified from extracts of Australian and Indopacific Zyzzya sponges [8,11]. Furthermore, two exotic pyrroloiminoquinones, atkamine and aleutianimine (Figure 3), have been isolated from Alaskan Latrunculia sp. [78] and Latrunculia austini [79], respectively. The structure and stereochemistry of atkamine were secured through chemical degradation, as well as comparison of experimental and TDDFT-simulated ECD spectra. Using a similar toolset incorporating DFT-simulated NMR spectra, the same group verified the structure of aleutianimine, providing a compelling example of the usefulness of computational approaches in natural product structure elucidation. Other unusual pyrroloiminoquinones are represented by the benzene derivative of discorhabdin C, isolated alongside various discorhabdins from an Alaskan deep-water Latrunculia sp. [12]. The same compound has been afforded semi-synthetically through dienol-benzene rearrangement [14] and is therefore considered to likely be an isolation artifact [12]. Veiutamine was isolated as a minor secondary metabolite alongside several common pyrroloiminoquinones from Fijian Z. fuliginosa and showed potent in vitro cytotoxicity in a panel of 25 cancer cell lines [80]. Veiutamine exhibits an unusual phenol-substituent directly bound to the pyrroloiminoquinone core and is to date the only known pyrroloiminoquinone with a C-6 p-oxy benzyl substituent.
In addition to true pyrroloiminoquinones, several closely related natural products have been isolated from marine sponges, often as minor secondary metabolites alongside makaluvamines, discorhabdins and tsitsikammamines (Figure 4). They include the secobatzellines from a Caribbean Strongylodesma sp. sponge [19] (reassigned from Batzella [81]); zyzzyanones from Australian Z. fuliginosa [45,82]; makaluvic acids from Micronesian Z. fuliginosa [40] and South African Strongylodesma aliwaliensis [83]; the oxazole-containing makaluvamine W from Tongan Strongylodesma tongaensis [53]; the structurally related citharoxazole from Mediterranean Latrunculia citharistae [84] and zyzzyamines from Papua New Guinean Z. fuliginosa [85].
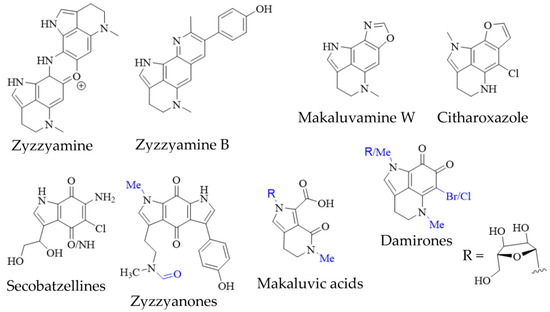
Figure 4. Pyrroloiminoquinone-related marine natural products from sponges. Variable substituents are shown in blue.
Pyrroloiminoquinone isolations commonly result in the recovery of ortho-quinone analogs of makaluvamines, named damirones [86] (Figure 4; some analogs are referred to as batzellines [87] and this structural archetype also encompasses makaluvamine O). As a result, such pyrrolo-ortho-quinones have been isolated from various makaluvamine- and isobatzelline-producing marine sponges including C. bellae (previously Latrunculia bellae [22,88]), S. aliwaliensis [48], Spongosorites sp. [67], Smenospongia aurea [89,90], Zyzzya spp. [7,8,39,41,42,45,46] and T. favus [49], in addition to the myxomycete D. iridis [26]. These compounds generally show greatly decreased cytotoxicity compared to e.g., makaluvamines that contain a pyrroloiminoquinone core [7,22]. Their formation from makaluvamines has been shown to be possible through, alkaline hydrolysis, lyophilization [91] and UV irradiation [92]. This, together with the fact that they in most cases have been isolated alongside makaluvamines, suggests that they may arise simply as degradation products. However, it can currently not be excluded that they may occupy a functional role in pyrroloiminoquinone biosynthesis.
2.3. Pyrroloiminoquinones and Related Pyrroloquinolines from Hydroids, Bacteria and Fungi
Pyrroloiminoquinone-related compounds have been isolated from several organisms unrelated to marine sponges or ascidians and this occurrence may hold important information to aid the identification of biosynthetic gene clusters or possibly even microbial symbionts responsible for or involved in pyrroloiminoquinone biosynthesis. Such organisms include the Australian marine hydroid Macrorynchia philippina which contains several cytotoxic macrophilones [27,28]. Some of these, exhibit a fully formed pyrroloiminoquinone core and have been shown to inhibit the conjugation of SUMO peptides to target proteins, eliciting greatly decreased levels of proteins involved in ERK signaling, while also exhibiting selective cytotoxicity in the NCI-60 anticancer panel [27,28]. In addition, related pyrroloquinoline alkaloids such as the lymphocyte kinase-inhibiting lymphostin [33] and the selectively cytotoxic ammosamides [34] are produced by marine-derived actinomycetes, Salinispora sp. and Streptomyces sp., respectively (Figure 5).
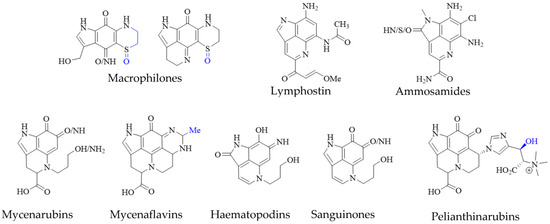
Figure 5. Pyrroloiminoquinone-related natural products from marine hydroids, marine soil-derived bacteria, and terrestrial fungi. Variable substituents are shown in blue.
While the vast majority of pyrroloiminoquinone structures have been reported from marine sources, terrestrial fungi of the genus Mycena have been shown to produce a range of pigments with clear structural relation to pyrroloiminoquinones (Figure 5). These comprise mycenarubins, mycenaflavins, haematopodins [30,32], sanguinones [29] and pelianthinarubins [31]. Some mycenaflavins have shown cytotoxic activity, while haematopodins and mycenarubins have been reported to exert antibiotic activity against soil bacteria [32]; however, compared with marine pyrroloiminoquinones, the biological activity of these fungal pyrroloquinolines has not been extensively investigated to date.
This entry is adapted from the peer-reviewed paper 10.3390/molecules27248724
This entry is offline, you can click here to edit this entry!