Hexavalent chromium is a highly soluble environmental contaminant. It is a widespread anthropogenic chromium species that is 100 times more toxic than trivalent chromium. Leather, chrome plating, coal mining and paint industries are the major sources of hexavalent chromium in water. Hexavalent chromium is widely recognised as a carcinogen and mutagen in humans and other animals. It is also responsible for multiorgan damage, such as kidney damage, liver failure, heart failure, skin disease and lung dysfunction. The fate of the toxicity of hexavalent chromium depends on its oxidation state. The reduction of Cr (VI) to Cr (III) is responsible for the generation of reactive oxygen species (ROS) and chromium intermediate species, such as Cr (V) and Cr (IV). Reactive oxygen species (ROS) are responsible for oxidative tissue damage and the disruption of cell organelles, such as mitochondria, DNA, RNA and protein molecules. Cr (VI)-induced oxidative stress can be neutralised by the antioxidant system in human and animal cells.
1. Introduction
Environmental contamination is defined as the elevated concentration of unwanted materials in air, water and soil beyond the permissible limit. It is also defined as an undesirable change in the natural environment that has harmful effects on both animals and plants [
1]. Heavy metals are naturally occurring elements having a higher density as compared to water [
2]. Chromium (Cr), arsenic (As), lead (Pb), iron (Fe), cadmium (Cd), nickel (Ni), mercury (Hg) and cobalt (Co) metallic ions are toxic even in low quantities [
3]. These heavy metals become lethal when their intake is in excess and when they are not metabolised by the body and accumulate in the intra- or extracellular space of body organs [
4,
5]. Heavy metals have been found in the body parts of fishes captured from the metal-contaminated aquatic system [
6]. Heavy metals enter the human body via the food chain [
7]. The effluent emanating from tanneries; battery-manufacturing units; glass, paint and metal plating industries; and pigment and steel productions cause heavy metal pollution in terrestrial and aquatic ecosystems [
8]. Coal has an important application in the energy sector and is considered a major energy source in India, China, Nepal, Pakistan and other countries. It is an easily available and inexpensive source of energy compared to other energy sources. These features make it a priority choice for several industries. During coal processing, a large volume of toxic substances is released as coal washery effluent (CWE), including heavy metal ions, such as Cd (II), Cr (VI) and Pb (II), which massively contaminate the natural environment [
9].
Cr (VI) has the seventh rank in the ATSDR [
10]. It is considered a priority carcinogen, which is generated from several anthropogenic activities, such as tanning of leathers, pigment and rubber production, paint manufacturing, production of anticorrosion agents and processing of coal [
11]. Cr (VI) is 100 times more toxic as compared to Cr (III). High solubility in water and a higher oxidative state make Cr (VI) more lethal. The Central Pollution Control Board (CPCB), India, has defined the maximum allowable concentration of Cr (VI) in the effluent of industrial units as 1.0–2.0 mg/L [
12]. Cr (VI) enters the human body and causes cancer, liver and kidney damage and difficulties in respiration [
13]. The World Health Organization (WHO) has recommended a permissible limit of up to 0.05 mg/L of Cr (VI) in drinking water [
14].
Several cell surface receptors are actively involved in the intake of Cr (VI) ions inside the cell [
15]. Cr (VI) ions in the intracellular space of the cell either bind with metallothionein proteins or are transformed into a less toxic form. Animal cells use intracellular Cr (VI) ions as electron acceptors or detoxify them by producing soluble enzymes [
15,
16]. Reactive oxygen species (ROS) are produced as an intermediate component during the reduction of Cr (VI) to Cr (III) [
17]. ROS are highly reactive, which damages cell organelles, and these Cr (VI)-mediated ROS can be neutralised by the antioxidant system [
18]. Antioxidants of the cell minimise Cr (VI) toxicity and protect the cell organelles from Cr (VI)-mediated oxidative damage [
15,
19]. Thus, antioxidant activity of the animal cell actively participates in the detoxification of Cr (VI) toxicity [
20].
2. Cr (VI)-Induced Oxidative Stress and Disruption of Cell Components
Cr (VI) is a well-known toxic agent, and its toxicity highly depends on the oxidation states and ionic species [
26]. Cr (VI) has higher solubility and reactivity as compared to Cr (III). It can easily cross the plasma membrane and enter the cell compartment through several cell surface receptor phosphate transporters and anion transporters [
27]. Cr (VI) is more hazardous due to its higher oxidative state and solubility [
28]. Cr (VI) has a negative impact on human health, which responsible for the damage of several organs, including the lungs, liver and kidneys [
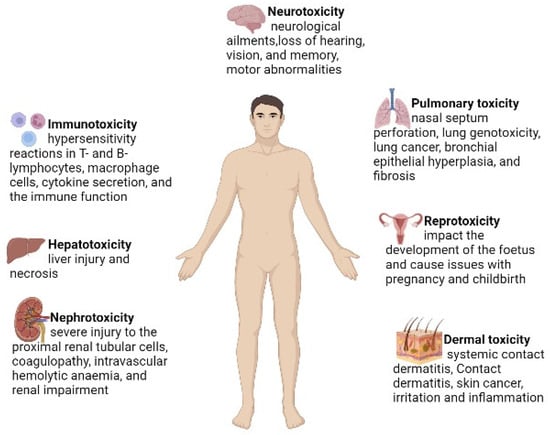
29]. The lethal effects of Cr (VI) are listed in
Figure 2.
Figure 2. The major hazardous effects of Cr (VI) on human health.
Several studies have reported that Cr (VI) causes carcinogenicity and multiple-organ damage, such as liver and cardiac failure and renal damage [
30]. Gumbleton and Nicholls (1988) reported that Cr (VI) induces kidney damage in rats after a sub-cutaneous injection of Cr (VI). Bagchi et al. [
31] reported that Cr (VI) induces hepatic mitochondrial, microsomal lipid peroxidation and increases lipid metabolites in the urine when Cr (VI) is administrated orally. Other important toxic effects of Cr (VI) are respiratory cancer, chromosomal abnormalities and DNA strand breaks [
32].
The reduction of Cr (VI) is considered a detoxification process when it occurs at a distance from the nucleus and other cell organelles or outside the cell. If Cr (VI) reduction occurs within the cell, it induces oxidative-mediated toxicity and damages cell organelles, and mutation in the DNA take place [
33]. In case Cr (VI) is converted into Cr (III) outside the cell, the reduced Cr (III) and other intermediate components are unable to transport into the cell compartment and hence a toxic effect is not observed [
34]. Cr (VI) passes through the cell membrane and enters the intracellular space and is subsequently reduced to Cr (III) [
35]. During the reduction process, ROS are generated, which cause cell toxicity. Studies suggest that Cr (VI) toxicity is mainly due to an increase in ROS production, which are produced by the Fenton reaction [
36,
37]. The generation of ROS in different cell lineages and their lethal impacts are mentioned in
Table 1.
Table 1. Cr (VI)-induced ROS generation and their lethal impacts.
| Cell Type |
Impact |
Citation |
| Cardiomyocytes of broiler |
Apoptosis-related genes Bax and p53 expression, mitochondrial malfunction and oxidative stress, myocardial apoptosis and autophagy |
[38] |
| Liver cell of zebrafish |
Downregulation of the Bcl2 gene and transcriptional activation of apoptosis-related p53, Bax, caspase 9 and caspase 3 genes, which changes the elemental composition of the liver |
[39] |
| Liver and kidney cells of Sprague-Dawley rats |
Dose- and time-dependent effects induced DNA damage due to increase in ROS levels |
[40] |
| Hepatocytes (HepG2) of humans |
Mitochondrial damage,
apoptosis, oxidative stress |
[41] |
| Lung epithelial cells of rats |
ROS-induced cell death and activation of the p53-related pathway |
[42] |
| Liver and kidney cells of carassius auratus |
Oxidative stress, genotoxicity and histopathology |
[43] |
| Bronchial epithelial cells (Beas-2B) of humans |
Cell transformation |
[44] |
| Liver and brain tissues of mice |
Oxidative stress and tissue damage |
[45] |
| Gill and kidney cells of Anguilla anguilla L. |
Genotoxicity at a higher concentration in gills and at all concentrations in kidneys |
[46] |
| Kidney cells of Wistar rats |
Apoptosis and autophagy, oxidative stress in kidneys, mitochondrial dynamics disorder via inhibiting the sirt/pgc-1a pathway |
[47] |
The toxicity of Cr (VI) is directly proportionate to its dosage and exposure time. Cr (VI) induces transcriptional upregulation of apoptosis-related genes, such as p53 and caspase (3 and 9), and downregulates genes involved in the antioxidant pathway [
38]. Long-term exposure to Cr (VI) causes higher oxidative stress in the cell and induces ROS-mediated cell death [
48].
Intracellular ROS cause DNA, RNA, protein and mitochondrial damage. The main mechanism by which Cr (VI) shows toxicity is through the disruption of transcriptional regulation, which makes it challenging to have normal gene expression pathways as complexation and DNA changes are more common in regions of active DNA replication and transcription [
49]. Several in vitro and in vivo studies have revealed that Cr (VI) induces oxidative stress by enhancing the production of ROS, which leads to deterioration of lipids and enzymes and DNA damage [
45]. In the cell, a cascade of cellular events occurs following Cr (VI)-induced oxidative stress, including enhanced generation of superoxide anions and hydroxyl radicals. In addition, Cr (VI)-induced oxidative stress is also responsible for increased lipid peroxidation, activation of protein kinase C, DNA fragmentation, alteration in gene expression, modulation of intracellular oxidised states and apoptotic cell death [
50]. Holmes et al. [
51] reported that Cr (VI) can inhibit the DNA replication and repair process. Bagchi et al. [
45] investigated the dose-dependent effects of Cr (VI) in female C57BL/6Ntac and p53-deficient C57BL/6TSG p53 mice. The authors observed the enhanced production of ROS and lipid peroxidation in the hepatic and brain tissues of female mice. Son et al. [
52] reported Cr (VI)-induced cell death of mouse skin epidermal cells by apoptosis or necrosis. Cr (VI)-induced cell death is a Cr (VI) dose-dependent phenomenon. Authors reported that several activities occur when cells are exposed to Cr (VI), such as cell shrinkage, migration of cells into the sub-G1 phase and increase in annexin V positively. The presence of Cr (VI) in culture media increases mitochondrial membrane depolarisation and caspase activation. Son et al. [
52] suggested that Cr (VI) induces mitochondrial-mediated and caspase-dependent apoptosis in mouse skin epidermal cells through activation of p53.
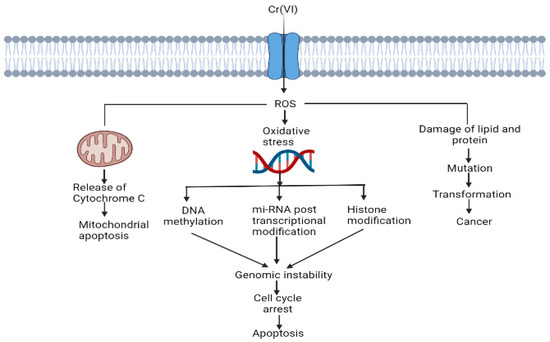
ROS-mediated disruption of cell organelles is represented in Figure 3.
Figure 3. Cr (VI)-induced ROS generation and their role in the disruption of cell organelles.
Cr (VI) alters histone modifications, miRNA expression and DNA methylation [
44]. Cr (VI) causes mitochondrial damage by oxidising Trx. For proper functioning of the cell, Trx should be in the reduced state, and the Trx system maintains a regular thiol redox balance and plays a role in cell survival [
53].
3. Antioxidants and Their Protective Role against Cr (VI) Toxicity
3.1. Enzymatic Antioxidants
3.1.1. Superoxide Dismutase (EC 1.15.1.1)
Superoxide dismutase (SOD) stubs out superoxide anions by altering them to peroxide, which is further wiped out by catalase or glutathione peroxidase [
57]. In aerobic respiration, a low concentration of superoxide is generated constantly [
54]. In mitochondria, the electron transport chain transfers four electrons to oxygen to form water. Sometimes, the electron transport chain leaks a single electron, which forms superoxide instead of water. Superoxide releases iron and reduces Fe (III) to Fe (II). Fe (II) reacts with H
2O
2 and generates hydroxyl radicals. SOD alters superoxide into H
2O
2 and O
2 [
54].
In human beings, in total, three forms of SOD have been recognised: (i) cytosolic Cu, Zn-SOD; (ii) extracellular-SOD; and (iii) mitochondrial SOD. Four classes of SOD have been recognised on the basis of cofactors, having either mononuclear Fe/Mn/Ni or binuclear Cu/Zn [
57,
58,
59].
SOD regulates inflammation, lipid metabolism, oxidation in cells and oxidative stress. SOD prevents lipid droplet formation, oxidation of lipoprotein in macrophages and lipid peroxidation [
54]. SOD is an effective antioxidant that reduces Cr (VI) chronicity and protects against oxidative-damage-related lethal effects.
3.2. Endogenous and Exogenous Antioxidants
Endogenous antioxidants are the by-products of human metabolism. An endogenous antioxidant may or may not be enzymatic [
74]. Superoxide dismutase is one of the enzymatic antioxidants involved in the first line of defence. Peroxiredoxins, catalase, glutathione reductase and glutathione peroxidase are some other important enzymatic endogenous antioxidants in the first line of defence [
74]. These enzymes neutralise hydrogen peroxide, yielding oxygen and water. Ceruloplasmin, ferritin, transferrin and albumin are examples of nonenzymatic molecules that participate in the body’s first line of defence and are preventive antioxidants found in the plasma [
74]. These proteins bind to transition metal ions to prevent the development of new reactive species. Additionally, metallothionein is also crucial for protecting against reactive species. It has a large number of –SH groups as the source of its main antioxidant effects [
74]. Many endogenous antioxidant mechanisms in living beings depend on exogenous antioxidants, such as polyphenols, carotenoids, vitamin E and vitamin C. To maintain redox homeostasis, endogenous and exogenous antioxidants work together synergistically [
75]. Examples include prevention of the lipid peroxidation process, which can cause damage to the cell membrane [
75]. Regeneration of vitamin E by vitamin C or glutathione can prevent the lipid peroxidation process, thus protecting cell from membrane damage [
75].
Many researchers agree that the synthesis of Cr (III) from the intracellular reduction of Cr (VI) and the subsequent oxidation of intracellular macromolecules (e.g., DNA) are essential. Indeed, it could be the cause of chromium’s carcinogenicity and mutagenicity [
76]. Intracellular reduction of Cr (VI) can be either enzymatic or non-enzymatic, depending on the tissues and other physiological parameters [
76]. The enzymatic reduction of Cr (VI) requires enzymatic antioxidant systems, such as nicotinamide adenine dinucleotide phosphate reductase, cytochrome c reductase, P450 cytochrome, catalase, glutathione S-transferase, glutathione peroxidase, glutathione reductase, superoxide dismutase, thioredoxin reductase, heme oxygenase biliverdin reductase and DT-diaphorase [
76].
3.3. Polyphenolic Antioxidants
3,4-Dihydroxybenzaldehyde (DHB), commonly known as protocatechualdehyde, is an antioxidant that protects the cell from Cr (VI) toxicity. Erythrocytes and lymphocytes were used for this investigation because DHB is both cyto- and geno-protective [
77]. The defence provided by DHB against inactivation by Cr (VI) may be a result of its inherent antioxidant characteristic, quenching the free radicals and ROS/RNS created by this metal ion, or it may be a result of DHB’s incorporation into the erythrocyte membrane, where it can affect the properties of enzymes/proteins [
78]. DHB prevents oxidative-stress-related cell death by preventing oxidative DNA damage and apoptosis [
35].
Ellagic acid is a well-known phenolic antioxidant and plays an important role against oxidative stress. Several studies were conducted on six groups of male Wistar rats to determine how renal damage caused by Cr (VI) works. It was found that ellagic acid shields rats treated with Cr (VI) that caused ROS stress in their renal tissues [
79]. Ellagic acid enhances glomerular filtration, tubular reabsorption and secretion processes. It also reverses histological alterations brought on by Cr (VI), lowers oxidative stress markers and boosts the activity of antioxidant enzymes [
80]. Ellagic acid may be able to chelate Cr
2+ and Cr
3+ intermediates, stopping cyclic Fenton/Haber–Weiss reactions since it chelates divalent cations [
81].
This entry is adapted from the peer-reviewed paper 10.3390/antiox11122375