Your browser does not fully support modern features. Please upgrade for a smoother experience.
Please note this is an old version of this entry, which may differ significantly from the current revision.
Subjects:
Biochemistry & Molecular Biology
Flavonoids are ubiquitous compounds of plants, produced by plants for growth and defense against all kinds of stress, including cold tolerance. More than 6000 different flavonoids have been identified, the primary sources of which are apples, red fruits, onions, citrus fruits, nuts, and beverages such as tea, coffee, beer, and red wine. These compounds, derived from phenol, are particularly interesting for their ability to cross the blood–brain barrier and for their multi-target activity. Several studies have described flavonoids to exhibit relevant biologic activities involving the neuronal antioxidants, as well as anti-amyloidogenic properties, acting as metal chelators, showing anti-inflammatory properties, and ameliorating cognition and neuroprotection.
- flavonoids
- neuroprotection
- quercetin
- myricetin
- epicatechin-3-gallate
1. Chemical Structure and Flavonoids Classification
From a chemical point of view, flavonoids consist of two benzene rings, called A and B, linked via a third pyranosic ring C. Flavonoids can be divided into a variety of subclasses that differ in terms of the structural characteristics of the B ring and the degree of hydroxylation and glycosylation of the third ring. Typically, ring B binds in position 2 on ring C, but can also bind in position 3 or 4. We can, therefore, distinguish the isoflavones in which the B ring binds in position 3 of the C ring and the neoflavonoids in which the B ring binds in position 4 of the C ring (Figure 2). The group of flavonoids in which the B ring is linked in position 2 of the C ring can be further divided into six subgroups, according to the structural peculiarities of the C ring: flavones, flavonols, flavanones, flavanonols, flavanols, catechins, and anthocyanins [58]. Finally, the flavonoids in which the C ring is open are called chalcones.
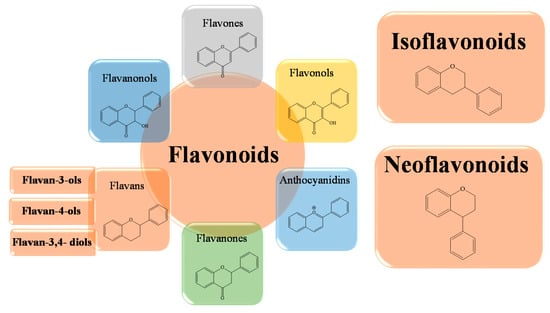
Figure 2. Schematic representation of the basic structures of flavonoid subclasses.
In general, since all flavonoids contain the same core scaffold, the functional differences between the various groups and subgroups are mainly due to the different substituent groups. These are weak polybasic acids of a polyphenolic nature, characterized by varying degrees of hydroxylation, methoxylation, glycosylation or glucuronidation, and this contributes to the great variety of biological properties of this large group of polyphenols [59,60]. In fact, a different side chain can significantly influence the activity of flavonoids on the same molecular target, and the total number of hydroxyl groups is important for the enhancement of antioxidant activity, free radical scavenging, and metal ion chelation [61,62,63,64].
2. Biological Activities of Flavonoids
Flavonoids are a wide group of secondary metabolites characterized by many interesting biological potentials, both in vitro and in vivo (such as anticancer, antioxidant, antiaging, anti-inflammatory, antimicrobial, and immunomodulatory activities, along with modulation of the activity of key metabolic enzymes, cytoprotective and cardioprotective potentials, and inhibition of cellular proliferation, for example) and, in the last decades, they have emerged as a promising agents for neuroprotection [58,65,66,67,68]. In a recent epidemiologic study, Shishtar et al. [69] analyzed long-term dietary flavonoid intake and risk of Alzheimer’s disease and related dementias in the Framingham Offspring Cohort, with a total of 5209 participants aged 28–62 years in the original cohort. The intake effects of six classes of dietary flavonoid (flavonols, flavones, flavanones, flavan-3-ols, anthocyanins, and flavonoid polymers) and the risk of Alzheimer’s disease and related dementias (ADRD) and Alzheimer’s disease (AD) alone were analyzed based on the data from the Framingham Heart Study Offspring Cohort. Participants were ADRD-free with a valid FFQ at baseline. Flavonoid intakes were updated at each exam in order to represent the cumulative average intake across the five exams, and were expressed as percentile categories of intake to handle their nonlinear relation with ADRD and AD. After multivariate and dietary adjustments, individuals with the highest intakes of flavonols, anthocyanins, and flavonoid polymers had a lower risk of ADRD relative to individuals with the lowest intakes. A similar trend was found also in AD for flavonols and anthocyanins, but not for flavonoid polymers, showing a direct correlation between higher long-term dietary intake of flavonoids and the risks of ADRD and AD onset in American adults.
Flavonoids are widely distributed in the plant kingdom, and are characterized by chemical structures with the presence of several substituents that allow them to assume particular activities and exert beneficial effects for the wellness of organisms, as well as for their potential for therapeutic utilization. This does not indicate that every type of flavonoid is able to show biological potential, but only that those with particular characteristics can be employed for specific roles. For instance, one of the well-known and best-studied activities of flavonoids is its antioxidant activity, which is linked to the number of hydroxyl groups on the B ring. Generally, a greater number of free hydroxyl groups corresponds to a greater scavenging effect, but their location in the skeleton of flavonoids is a crucial structural element. These hydroxyl groups, through the donation of hydrogen atoms and electrons to radical species, favor the repair of the damage caused by ROS and reactive nitrogen species (RNS), reducing their degree of reactivity [70,71]. This mechanism leads to the generation of a relatively more stable flavonoid radical, which significantly reduces the oxidative stress triggered by the interrupting free radicals. At the level of the cell membrane, there is a chain reaction of propagation of peroxylic radicals between the molecules of polyunsaturated fatty acids and other intermediates [72,73]. The scavenger action of flavonoids on ROS and RNS is important because the ROS/RNS balance is directly connected with the redox state of the cell, which is also influenced by the presence of metal ions. In addition to the direct action on free radicals, experimental data demonstrate specific chelating properties of flavonoids against transition metals, mainly iron and copper ions [74,75,76]. Ferrous and/or copper ions represent a danger because they tend to react through the Fenton reaction, with hydrogen peroxide generating hydroxyl radicals, a very reactive species which rapidly oxidizes surrounding molecules, triggering the oxidative stress cascade. In a second antioxidant mechanism, flavonoids do not act directly on ROS, but “indirectly” interact with some proteins involved in the gene expression regulation pathway. They upregulate the endogenous antioxidant capacity of the cell, and inhibit others involved in redox balance and inflammatory processes, such as cyclooxygenase, lipoxygenase, xanthine oxidase, NADH oxidase, and myeloperoxidase [83,84,85,86,87,88,89]. Therefore, the mechanisms we have described suggest not only a direct involvement of the flavonoid molecule, but also of metabolites that result from its oxidation and the formation of flavonoid–metal complexes [90].
3. Flavonoids in Neurodegeneration
The ability of flavonoids to cross the blood–brain barrier suggests that these compounds can feasibly have a direct effect on the brain. Numerous studies have documented the bioactivity of flavonoids against neurodegenerative disorders such as AD, Parkinson’s, Huntington’s, and other neurological disorders [51,58,64,65,66,67,68,69,91,92,93]. Regarding AD treatment, there is still no significantly efficient drug that can reduce the progression or improve the outcome of the disease [94,95]. The search for natural substances for the treatment of AD is considered key to brain health, because these compounds are often easily isolated and possess well-documented biomechanisms and safety profiles [96,97]. In addition, their abundance in vegetables and fruits has made them a major part of the human diet.
4. Potential Role of Flavonoids in AD Therapy
Flavonoids, including epicatechin-3-gallate, gossypetin, naringerin, quercetin, and myricetin are reported to block β-amyloid and Tau aggregation, scavenge free radicals, and sequester metal ions at clinically low concentrations [98,99]. In order to better understand flavonoids’ role in AD treatment, the pharmacological effects of some compounds from the six subclasses are described (see Figure 3).
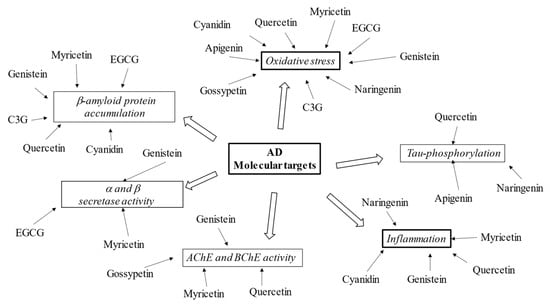
Figure 3. Schematic effects of described flavonoids on molecular targets of AD.
Quercetin is a polyhydroxyflavonoid that belongs to the subclass of flavonols. Its chemical name is 3,3,4,5,7-pentahydroxyflavone, and the molecule contains five -OH groups, in positions 3, 3′, 5, 7 and 4′, which are crucial for potential biochemical–pharmacological activities. Quercetin is a natural antioxidant, widely used in healthcare for its beneficial role. Quercetin is found in flowers and fruits of edible plants; onions, apples, cherries, berries, asparagus, and red leaf lettuce have the highest levels, while tomatoes, peas, and broccoli have lower levels [92,93]. Experimental studies have shown the existence of an inverse correlation between dietary quercetin intake and risk of senile dementia. Specifically, experimental studies have identified various quercetin targets of neuronal protection that contribute to reducing the main neuronal lesions present in the brains of AD patients. Among them are hyperphosphorylation of the Tau protein, the deposition of beta amyloid, oxidative stress, inflammation, and apoptotic processes [100,101].
More specifically, Tau phosphorylation is under the control of several distinct kinases, such as Erk, Akt, p38, AMP activated protein kinase (AMPK), glycogen synthase kinase 3 beta (GSK3β), cyclin-dependent kinase 5 (cdk5), and protein phosphatase 2A (PP2A) [102,103]. Through molecular dynamic simulation studies carried out according to the molecular docking results, Zu et al. identified MAPK as core target of quercetin. Jiang et al. demonstrated the anti-apoptotic role of quercetin via MAPKs and PI3K/Akt/GSK3ß signaling pathways by preincubating HT22 cells with 5 μmol/L of the drug for 12 h [103,104]. MAPK is a heterotrimeric Ser/Thr protein kinase; its activation contributes to the hyperphosphorylation of Tau in neurons. It also checks Aβ metabolism, and is involved in cell proliferation, apoptosis, and inflammatory responses [105]. Thus, MAPK pathway inhibition can significantly improve synaptic plasticity, memory, and cognitive functions, and can be considered a valid target against AD progression [106]. GSK3β, a Ser/Thr kinase that connects numerous signaling pathways in the cell, including inflammatory responses, is another strategic target for neuroprotection. GSK3β connects numerous signaling pathways in the cell. It is a downstream enzyme of the PI3K/Akt signaling pathway, and is considered the main factor responsible for the phosphorylation of the Tau protein in AD [107]. Quercetin acts on GSK3β, decreasing the kinase activity, and therefore, quercetin indirectly has an anti-hyperphosphorylation of Tau protein action that helps to strengthen its neuroprotective effects [104]. In AD, the decrease in the expression of PGC-1α is one of the causes related to the progression of the disease, as it is linked to the increase in the generation of the peptide Aβ [109,110]. In addition, the quercetin action on p53 affects the oxidative stress decrease, because p53 is a significant activator of the ROS-mediated apoptotic pathway [111,112]. A total of 50 μM quercetin, after 4 h of HepG2 cells incubation, has been shown to restore cellular redox homeostasis through its significant scavenger activity against ROS. Feeding the C57BL/6J mice with a 1% quercetin diet for 20 weeks increased the level of glutathione (GSH) and the expression of certain antioxidant enzymes, including SOD, catalase (CAT), and glutathione peroxidase (GPx), in hippocampal neurons [113,114,115,116].
In the brain, BChE also appears to play a role in the transformation of dangerous amyloid plaques to the pathogenic structures present in dementia and AD [123,124]. In Figure 4, a schematic representation of the main activities of quercetin is depicted.
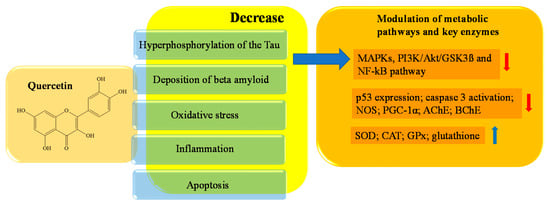
Figure 4. Schematic effects of quercetin on molecular targets of AD.
Naringenin is the aglycon of naringin, and belongs to the subclass of flavanones. It is abundant in citrus fruits (especially grapefruits, to which it gives the characteristic bitter taste), as well as vegetables, and especially in grapes, tomatoes, and cherries. Naringenin can be found in two forms. One is characterized by a bond with a sugar on C7, and one derives from the action of specific enzymes which are able to cleave this glycosidic bond by releasing the aglycone [125,126]. Both forms have antioxidant activity, but naringenin has a more powerful scavenging action than its precursor naringin, because the presence of sugar in the latter causes a steric hindrance that impairs activity [127]. Naringenin (400 mM) improved learning and spatial memory in PC12 cells of rats with AD through the regulation of the PI3K/Akt/GSK-3K pathway and by reducing the hyper-phosphorylation of TAU. The intracellular mechanism that allows this neuroprotective action is related to the inhibition of caspase 3 activity, the activation of PI3K/Akt, and the modulation of the signaling pathway GSK3β, which plays a crucial role in neuronal survival [128,129,130]. The inhibition of caspase 3 also affects programmed cell death; in fact, the block of neuroapoptosis is also caused by the decreased levels of malondialdehyde and hippocampal nitrite found in socially defeated rat pups treated with naringenin (50–100 mM) [131]. Naringenin significantly regulates the (NF-kB) signaling pathway, which is implicated in inflammatory processes, and decreases tumor necrosis factor-α (TNF-α) as well as interleukin (IL)-6 and IL-1β [131]. The reduced expression of NF-kB is induced by naringenin through a significant decrease in phosphorylation and nuclear translocation of P65, a subunit of NF-kB, as well as by an increase in sirtuin 1 (SIRT1) levels in the hippocampus [132,133,134,135]. The inflammatory pathway is also repressed by the interaction of naringenin with other molecular targets, including inducible nitric oxide synthase (iNOS) and cyclooxygenase 2 (COX-2) e MAPK. iNOS and COX-2 are both inhibited in a concentration-dependent manner, while Zhang et al. demonstrated that naringenin (100 mM) inhibited MAPK signaling pathway activation by suppressing the phosphorylation of JNK and ERK1/2 in BV-2 cells [136]. Numerous studies have reported the antioxidant properties of polyphenols; it has been shown that GSH activity, one of the most efficient endogenous antioxidants, is improved by naringenin. Additionally, an increase in SOD, CAT, GPx, and glutathione reductase (GR) activity, in addition to a decrease in hydrogen peroxide (H2O2) and protein carbonyls levels, have been demonstrated [137,138,139,140]. In Figure 5, a schematic representation of the main activities of naringenin is depicted.
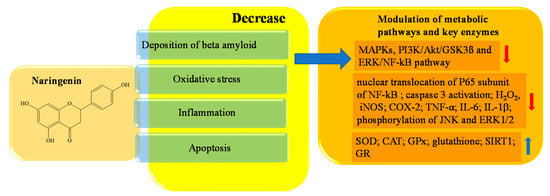
Figure 5. Schematic effects of naringenin on molecular targets of AD.
Epigallocatechin-3-gallate (EGCG), an ester of epigallocatechin and gallic acid, is the main bioactive polyphenol found in solid green tea extract [141,142]. EGCG has been reported to bypass the blood–brain barrier (BBB) and exert potent neuroprotective properties against AD, in a wide range of cell models [143]. EGCG enhances cholinergic neurotransmission through inhibition of AChE and BChE. ACh content was significantly elevated in a dose-dependent manner, which ultimately led to an improvement in the learning and memory function of AD rats [152]. In Figure 6 a schematic representation of the main activity of epigallocatechin-3-gallate is depicted.
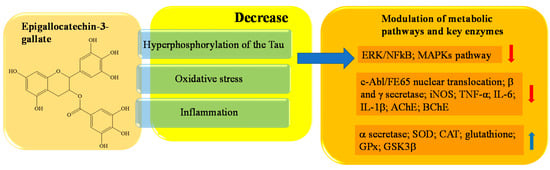
Figure 6. Schematic effects of epigallocatechin-3-gallate on molecular targets of AD.
Myricetin is a natural flavanol widely distributed in several vegetables and fruits, mainly including blackcurrant teas, red wines, and medical herbs [153,154,155,156]. Myricetin, mainly in the form of glycoside (O-glycosides), is also known as hydroxy quercetin because of its quercetin-like structure [157,158], from which it differs by one extra hydroxyl at the 5′-OH of the B ring. The compound has a wide range of beneficial effects on human health, including antihypertensive, antiallergic, analgesic, anti-inflammatory, immunomodulatory, antiplatelets, and aggregation activities [158,159,160]. Myricetin inhibits lipopolysaccharide (LPS)-induced neuroinflammation, as it reduces the levels of proinflammatory mediators, including IL, NF-kB, TNFα, iNOS and COX2, in the microglia BV2 cell line. [178]. Finally, other myricetin-like polyphenols, including gossypetin, also has inhibitory abilities against AChE [179,180]. In Figure 7, a schematic representation of the main activity of myricetin is depicted.
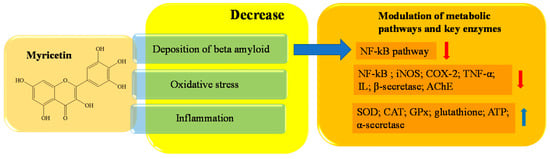
Figure 7. Schematic effects of myricetin on molecular targets of AD.
Gossypetin (3,5,7,8,3′,4′-hexahydroxy flavone) is a flavonol isolated from the flowers and the calyx of Hibiscus sabdariffa. Gossypetin has been shown to exert antioxidant, antimutagenic, antimicrobial, and anti-atherosclerotic activities [181,182,183,184]. Chen et al. [183], in murine macrophage cell line J774A.1, demonstrated that gossypetin (1–1000 μM) has inhibitory effects on both lipid peroxidation and lipoprotein oxidation, attenuating the formation of foam cells and lipid accumulation through PPAR pathways. The drug improves cholesterol removal from macrophages and delays atherosclerosis [183]. In addition, Lin et al. [184] demonstrated that gossypetin (0.1–0.5 μM) has inhibitory effects on abnormal vascular smooth muscle cell proliferation and migration, which could lead to the containment of atherosclerosis and other cardiovascular illnesses [184]. Inhibition of AChE and BChE activity, key enzymes in brain-related disorders, further emphasizes the therapeutic benefit of gossypetin for the treatment of AD [180,185].
Genistein (4′,5′,7-trihydroxyisoflavone) is an isoflavone distributed in several vegetables such as legumes, green peas, and peanuts, and is predominantly extracted from the Glycine max soybean [186,187]. Several researches point out genistein potential therapeutic role to delay the onset of Alzheimer’s dementia through the improve of cognitive function and synapse development [188]. In this context, Safahani et al. demonstrated that genistein supplementation modulates dopaminergic and cholinergic function, helping with memory recovery and neuroprotection in rats [189]. Genistein (10 mg/kg) protects against AD progression by reducing the production and deposition of Aβ aggregates, as well as the hyperphosphorylation of the Tau protein, in rat models [188,190,191]. Genistein protects against AD progression by reducing the generation and aggregation of Aβ. Seong et al. showed that the inhibitory activity (%) of genistein against the extent of Aβ25–35 self-aggregation, after 24 h of incubation, decreased by 34.90% when co-treated with 100 µM genistein [192]. In rat hippocampal neurons, the drug (0.375 µg/mL) downregulated presenilin levels, increased α secretase while decreasing β secretase and BACE1 activity, and, last but not least, modulated the PPARγ receptor to upregulate ApoE production [193,194,195,196]. In vitro genistein (0.391 mM) has been effectively proved to reduce oxidative stress, due to its high antioxidant power and potent ROS scavenger ability [197].
The antioxidant effects of genistein are associated with AMPK activation and the drug’s binding with estrogen receptor α (ERα), both of which promote the expression of antioxidant enzymes such as SOD, CAT, and GPx [198,199,200]. Moreover, in RAW 264.7 cell model, genistein (20 mM) prevents neuro-inflammation by regulating gene transcription of cytokines, such as TNFα, IL-1β, IL-6, and IL-12 [201,202]. Finally, Fang et al., 2014, demonstrated the inhibitory activity of AChE by two genistein derivatives, highlighting the therapeutic potentiality of this isoflavone [203].
Apigenin (4′,5,7-trihydroxyflavone) formally belongs to the flavone subclass, and is widely distributed in the plant kingdom, present principally in chamomile flowers. and in lower concentrations in vegetables, citrus fruits, herbs, and plant-based beverages (tea, beer, and wine) [204]. In a rat model of AD, apigenin (50 mg/kg) significantly reduced the hyperphosphorylation of tau levels in the hippocampus, decreasing the expression of GSK-3β, suppressing BACE1 expression, and supporting an antiamioloidogenic activity [205]. The drug (25 µM in human THP-1 monotypic cells) inhibits the production of IL-6 and IL-1β by modulating the MAPK/ERK and PI3-K/Akt signal transduction pathways associated with neuronal survival blocking [206,207,208]. Apigenin decreases ROS levels and significantly increases GSH levels, improving the cellular antioxidant defense system [209,210].
Cyanidin 3-O-glucoside belongs to the anthocyanins class; it is present in plants such as berries and soybean fruits, where it is responsible for the red, purple, and blue pigments. Cyanidin has been reported to act as neuroprotector in several disorders, such as AD, Parkinson’s disease, and multiple sclerosis [211,212,213,214,215]. The anti-AD effect is evidenced in the inhibition of Aβ amyloid accumulation. Cyanidin could directly interact with Aβ peptide through hydrogen bonds responsible for Aβ aggregation and fibrils formation [216]. In addition, Thummayot et al. (2014, 2016) demonstrated that, in human neuroblastoma (SK-N-SH) cells, cyanidin (20 μM) decreases Aβ-induced apoptosis of SK-N-SH cells by decreasing the expression levels of several proteins. This facilitates the release of the pro-apoptotic factors required for caspase cascade activation [217,218]. The neuroprotective effects of cyanidin may also be mediated via inhibition of oxidative stress and pro-inflammatory cytokines release. The compound is a scavenger of radical activity, a strong inhibitor of intracellular ROS generation, and an enhancer of the cellular antioxidant system. It has been demonstrated that pretreatment of the cells with cyanidin improved the expression of antioxidant enzymes, namely SOD, CAT, and GPx [219,220,221]. Moreover, Kaewmool et al. showed that, in LPS-stimulated BV2 microglia, cyanidin (2.5–10 mM) inhibits the signaling pathways NF-κB and p38, while MAPK suppresses the production of interleukin-1β (IL-1β) and interleukin-6 (IL-6) and subregulates the gene expressions of iNOS and COX-2 in BV2 cells [222].
This entry is adapted from the peer-reviewed paper 10.3390/ijms232314835
This entry is offline, you can click here to edit this entry!