Your browser does not fully support modern features. Please upgrade for a smoother experience.
Please note this is an old version of this entry, which may differ significantly from the current revision.
Subjects:
Cardiac & Cardiovascular Systems
|
Oncology
Angiogenesis is the process of new blood vessel formation through the migration, growth, and differentiation of endothelial cells.
- cancer
- cardiovascular disease
- miRNA
1. Introduction
Cancer and cardiovascular disease are the leading causes of death across the globe accounting for one in six deaths [1] and 32% of all deaths worldwide [2], respectively, according to World Health Organization (WHO). Both cancer and cardiovascular disease are the umbrella terms commonly used to describe several disease etiologies. Each etiology of cancer and cardiovascular disease (e.g., lung cancer and ischemic heart disease, respectively) has its own distinct cause and progression pattern. However, recent research suggests that many aspects of cancer and cardiovascular disease are similar in terms of pathogenesis [3,4,5], leading to the development of specific field of study called cardio-oncology [6,7]. For example, both diseases involve dysregulated functionalities in vasculature, where abnormal vasculature (called, tumor vasculature [8]) occurs in cancer, while coronary artery disease is a type of cardiovascular disease caused by the narrowing or blockage of coronary arteries [9]. Another example is the involvement of immune responses, where prolonged or chronic inflammation is a hallmark of cancer [10,11,12] as well as cardiovascular disease [13,14,15]. The activation of immune responses often leads to the deposition of excessive extracellular matrices [16,17], which are another hallmark of cancer [17] and cardiac fibrosis as the end-stage of heart failure [18].
MicroRNAs (miRNAs) are evolutionary-conserved, regulatory short [~22 nucleotides (nt)] non-protein-coding RNAs that function as translational inhibitors by binding to the 3′-untranslated regions (3′-UTRs) of messenger RNAs (mRNAs) [19,20]. As one miRNA is predicted to bind hundreds of mRNAs due to its very short seed sequence (~6 nt) [21,22,23], it is speculated and experimentally shown for some miRNAs to regulate cascades of signaling pathways and their downstream targets. Due to their versatilities, dysregulation in miRNAs is linked to a variety of diseases, including cancer [24,25] and cardiovascular disease [26,27,28]. As the regulatory importance of miRNAs is experimentally proven, the therapeutic silencing of miRNAs is being explored [29,30,31,32,33,34]. However, due to their biodistributions (e.g., including their presence in the circulation [35,36,37]) and the presence of many target mRNAs for one miRNA, the precise mechanistic elucidation of each miRNA is urgently needed to advance into clinics. Since a specific miRNA is highly dependent on which target mRNAs are present in a specific biological context, it must be taken into consideration that the same miRNA can yield different biological outcomes depending on the specific cell or tissue [38]. This is especially important when considering miRNAs as potential therapeutic targets.
As cancer and cardiovascular disease share several aspects of disease causes and progressions, it is no surprise that many miRNAs are shown to be involved in pathogeneses of both cancer and cardiovascular disease. Because the heart is the least likely organ to harbor tumor growth [39], the communication between researchers working in miRNAs for either cancer or cardiovascular biology is scarce, although many miRNAs are found to be dysregulated in both diseases.
2. Angiogenesis: AngiomiRs
Angiogenesis is the process of new blood vessel formation through the migration, growth, and differentiation of endothelial cells [57,58]. In cancer, angiogenesis allows for a tumor to grow as new vessels provide nutrients and oxygen to malignant cells [59,60]. In cardiovascular disease, therapeutic angiogenesis aims to provide the blood flow to the ischemic heart tissue [61,62]. Thus, in both diseases, angiogenesis is an important therapeutic target, although the opposite effects are observed. During angiogenesis, several miRNAs are functionally involved, which has created a specific term to describe these angiogenesis-related miRNAs called, angiomiRs (Figure 2). AngiomiRs include miR-15/16, miR-17~92 cluster, miR-18a, miR-19, miR-21, miR-23b, miR-27a/b, miR-29b, miR-30, miR-34a, miR-57, miR-125b, miR-126, miR-128, miR-143, miR-145, miR-155, miR-192, miR-194, miR-199a, miR-200 family, miR-204, miR-210, miR-217, miR-296, miR-378, miR-484, miR-494, miR-497, miR-542-3p, miR-573, miR-642, and let-7b [63,64], which some are discussed below.
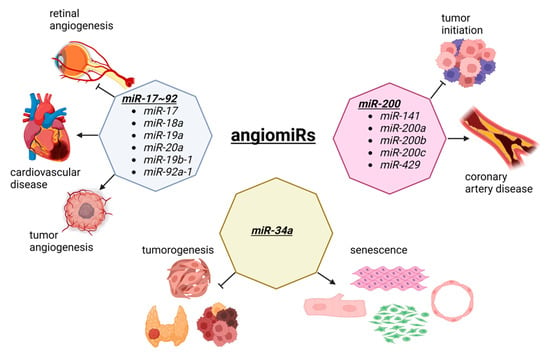
Figure 2. The dual role of angiomiRs in cancer and cardiac pathophysiology. The miR-17~92 cluster is involved in tumorigenesis and tumor vascularization. This cluster is also involved in retinal angiogenesis and the progression of cardiovascular disease. The members of the miR-200 family prevent the tumor initiation and malignant transformation, although they are upregulated in coronary artery disease. MiR-34a is a tumor suppressor involved in the development of thyroid cancer, head and neck squamous cell carcinoma, and cancer stem cells division. The overexpression of miR-34a suppress the proliferation and induces senescence in cardiomyocytes, fibroblasts, smooth muscle, and endothelial cells, by inhibiting sirtuin 1 (SIRT1). Figure created with BioRender.com, accessed on 24 October 2022.
The miR-17~92 cluster was first reported in tumorigenesis [65] and is one of the most well-studied miRNA clusters [66,67]. By crossing miR-17~92 floxed mice with an inducible vascular endothelial cell specific Cre driver (Cdh5-cre/ERT2), Chamorro-Jorganes et al. demonstrated that retinal angiogenesis was reduced during the development of these mice [68]. Furthermore, the vascular endothelial growth factor (VEGF)-induced ear and tumor angiogenesis were reduced, suggesting that VEGF regulates miR-17~92 cluster expression leading to the regulation of angiogenesis. The involvement of the miR-17~92 cluster is well documented in various diseases, including cardiovascular disease [69,70]. Since the miR-17~92 cluster consists of miR-17, miR-18a, miR-19a, miR-20a, miR-19b-1, and miR-92a-1, each miRNA in this cluster is also shown to be important for angiogenesis, including tumorigenesis and cardiovascular disease. For example, miR-92a is dysregulated in many forms of cancer, suggesting it is a potential diagnostic biomarker as well as a therapeutic target [71]. In the cardiovascular system, Bonauer et al. demonstrated that overexpression of miR-92a in endothelial cells inhibited angiogenesis in murine models of limb ischemia and myocardial infarction, while the silencing of miR-92a via antagomiR resulted in enhanced angiogensis and the functional recovery of the damaged tissues in murine disease models, suggesting miR-92a as a potential therapeutic target for ischemia diseases [72].
The miR-200 family is another well studied miRNA family that includes miR-141, miR-200a, miR-200b, miR-200c, and miR-429 [73]. In cancer, the miR-200 family is shown to play functional roles in cell malignant transformation and preventing tumor initiation [74]. By profiling epicardial adipose tissue from coronary artery disease (CAD) patients and non-CAD atherosclerotic patients, Zhang et al. demonstrated that the expressions of miR-141-3p, miR-200b, miR-200c-3p, and miR-429 are up-regulated in CAD patients compared to non-CAD patients [75]. By performing a series of experiments in vitro, the authors demonstrated that the overexpression of miR-200b-3p in human umbilical vein endothelial cells (HUVECs) resulted in increased apoptosis under oxidative stress. Mechanistically, miR-300b-3p targets histone deacetylase 4 (HDAC4) as the overexpression of HDAC4 reduced the increased apoptosis induced by inhibiting miR-200b-3p, suggesting that miR-200b-3p is a potential therapeutic target for atherosclerosis.
MiR-34a is a tumor suppressor and considered as a diagnostic and prognostic biomarker as well as a therapeutic target in various cancers, including head and neck squamous cell carcinoma, thyroid cancer, and cancer stem cells [76,77]. Interestingly, the expression of miR-34a is increased in senescent HUVECs and in the heart and spleen of older mice [78]. When overexpressed, miR-34a suppressed cell cycle and proliferation by inhibiting sirtuin 1 (SIRT1). Because ageing is a hot topic to be investigated, subsequent research shows the functional importance of miR-34a in cell types other than endothelial cells in the heart, including in cardiomyocytes [79,80], fibroblasts [81], and smooth muscle cells [82,83]. This is not an isolated case as many other angiomiRs (and other miRNAs) are expressed rather ubiquitously, suggesting that examining miRNAs as a common mechanism of action for cardio-oncology is not a big surprise.
This entry is adapted from the peer-reviewed paper 10.3390/cells11223551
This entry is offline, you can click here to edit this entry!