2. Clinical Phenotypes of CMA
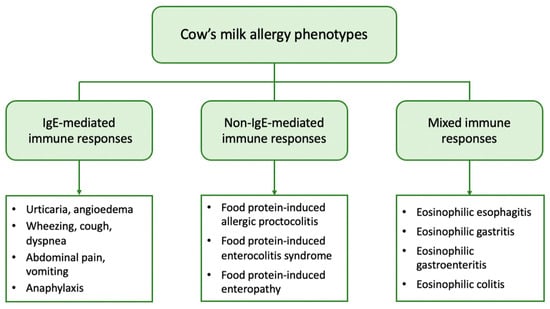
From a clinical point of view, patients with CMA may present with a variety of symptoms linked to the immune mechanism underlying the allergic reaction. Based on the mechanism involved, CMA can be further classified into (a) IgE-mediated, which is caused by antibodies against milk proteins belonging to IgE, (b) non-IgE (or cell-mediated), where the cellular immune system, and especially T-cells, are responsible for the allergic reaction, and (c) mixed type when both IgE and immune cells are involved [
18].
IgE-mediated CMA is the most common form and accounts for approximately 60% of all CM-induced allergic reactions [
19]. It is caused by the production of specific IgE against CM proteins that bind to high-affinity IgE receptors (FcƐRI) on basophils and mast cells. Upon exposure, CM proteins are recognized by two or more specific IgEs that bind to FcƐRI with subsequent cross-linking of the receptor and activation of mast cells to release mediators such as histamine, tryptase, and platelet-activating factor [
20]. These mediators cause vasodilation and elicit acute symptoms in the skin, gastrointestinal, respiratory, and cardiovascular systems. IgE-mediated reactions typically occur immediately after CM ingestion, or within 1 to 2 h, and may present as acute urticaria, angioedema, cough, wheezing, dyspnea, abdominal pain, vomiting, and hypotension. In severe cases, anaphylaxis, a systemic reaction that can be fatal, can also occur [
18]. The diagnosis of IgE-mediated CMA is based on the combination of a compatible medical history and evidence of CM sensitization, i.e., the presence of specific IgE in skin mast cells (skin prick tests, SPTs) and/or in serum [
21].
The non-IgE-mediated types of CMA have been far less studied. They encompass a wide range of disorders including food protein-induced allergic proctocolitis (FPIAP), food protein-induced enterocolitis syndrome (FPIES), and food protein-induced enteropathy (FPE). These disorders affect different segments of the gastrointestinal tract (GIT) and are distinguished by the delayed onset of symptoms, which can range from 2 h to several days after CM ingestion. They are caused by different immune cell-mediated mechanisms but have in common that they all lead to inflammation of the GIT. In these types of CMA, there is no evidence of CM sensitization, as no circulating specific IgE is detected [
22,
23]. However, a localized IgE response to the gut has been described [
24].
FPIAP is one of the most common allergic diseases in infancy. It is characterized by the presence of mucus and blood in the stools of an otherwise healthy infant. Unlike other types of CMA, this clinical phenotype most commonly affects breastfed infants. It usually occurs in infants up to 3 months of age, and most of them develop tolerance within the first year of life. Diagnosis is based on clinical manifestations [
23].
FPIES occurs less frequently. In two large prospective cohort studies from Israel and Spain, the cumulative incidence of CM-FPIES was 0.34% and 0.35%, respectively, in children during the first two to three years of life [
25,
26]. It mainly affects infants younger than nine months and is differentiated into acute and chronic FPIES. Infants with acute FPIES appear with multiple vomitings within a few hours of CM intake, which may be accompanied by pallor, lethargy, and diarrhea. In contrast, infants with chronic FPIES exhibit more chronic symptoms, such as vomiting, chronic diarrhea, and inadequate growth. Diagnosis in both cases is based on clinical criteria [
27].
FPE is considered relatively uncommon, although its prevalence has not been thoroughly investigated. This disorder affects the small intestine and presents with malabsorption symptoms, such as chronic diarrhea, bloating, flatulence, anemia, and failure to thrive [
28].
Mixed types of FA comprise a group of diseases generally referred to as eosinophilic gastrointestinal disorders (EGID). They are triggered by complex immune mechanisms with only partial involvement of IgE, leading to pathological eosinophilic infiltration of different parts of the GIT. Symptoms have a delayed onset and depend on the organs affected, as well as the extent of eosinophilic infiltration [
29]. Eosinophilic esophagitis (EoE) is the most common disease, with CM and wheat being the main culprits. EoE in children presents with feeding difficulties, gagging, vomiting, and food refusal due to gastroesophageal dysfunction, and the diagnosis is based on endoscopic findings [
22].
Atopic dermatitis (AD) is the most common chronic skin disease in childhood with an estimated prevalence of up to 20% [
30]. It is a complex disease that is characterized by impaired skin barrier function due to various genetic, environmental, and immunologic factors. AD is usually the initial manifestation of the “atopic march” and predisposes to other allergic disorders such as FA, asthma, and allergic rhinitis [
31]. Approximately one-third of children with moderate to severe AD develop FA, with CM being one of the main allergens [
32]. However, the role of CM in the development of AD remains controversial, with many authors supporting the idea that CMA is a consequence of AD rather than AD being an expression or clinical phenotype of CMA [
33]. In contrast, there are limited reports in the literature of isolated delayed-type eczematous reactions that typically occur within 6 to 48 h after food ingestion with eczematous flare-ups at AD-predisposed sites, suggesting a non-IgE-mediated pattern [
32].
Lactose intolerance (LI) is a common gastrointestinal disorder that is caused by lactase deficiency and the inability to digest and absorb dietary lactose [
34]. Lactose is the major carbohydrate in human and mammalian milk and is absorbed in the small intestine after first being hydrolyzed into D-glucose and D-galactose by the enzyme lactase [
35]. Lactase expression is genetically programmed to decline gradually after weaning in about 70% of the world’s population [
36]. As a result, lactase levels progressively decrease over time and its deficiency leads to lactose malabsorption and subsequent microbial fermentation of unabsorbed lactose in the colon [
34]. Fermentation products, especially gasses, affect gastrointestinal function and may lead to the development of clinical symptoms, such as abdominal pain, flatulence, and diarrhea after consuming milk and dairy products. Despite similarities in clinical presentation with CMA, LI does not affect the immune system and is therefore not considered a FA. Moreover, it rarely manifests clinically in children younger than five years of age [
37] and can be diagnosed by a variety of diagnostic methods, such as the hydrogen breath test [
38].
3. Gut Microbiome and the Factors Influencing Its Development in Early Life
The GIT is colonized by a wide variety of microorganisms, mainly bacteria, which perform substantial metabolic, immunological, and gut protective functions. The structure of these complex microbial communities varies along the GIT and shows greater diversity in the colon where the bacterial density reaches 10
12 colony-forming units (CFU)/mL [
39]. Despite the large diversity, only four major phyla, Firmicutes, Bacteroidetes, Actinobacteria, and Proteobacteria, are represented in the human gut [
40]. However, the exact taxonomic composition depends on many host-related factors, including genetic variation, age, diet, and geographic location, and thus varies significantly among healthy individuals [
41].
The gut microbiome is acquired at birth and undergoes dynamic changes during the first years of life. In newborns it is initially dominated by Proteobacteria (e.g.,
Escherichia) and Actinobacteria (e.g.,
Bifidobacterium). Its composition is constantly changing and increasing in diversity throughout infancy and is largely mature by the age of two to three years. It then tends to acquire a more stable composition dominated by Firmicutes and Bacteroidetes and is characterized by distinct functions [
42]. The ratio of Enterobacteriaceae to Bacteroidaceae, referred to as the E/B ratio, reflects maturation in the adult-type microbiome and decreases with age [
43].
However, recent evidence suggests that the colonization of the gut may begin in utero, contrary to the widespread theory that the fetal environment is sterile [
44,
45,
46]. Hence, the first 1000 days of life, or the time period from conception to two years of age, is considered the critical window for shaping the microbiome and regulating the immune system [
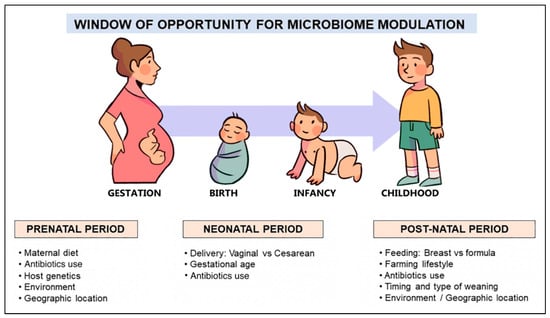
47]. Mode of delivery, gestational age, breastfeeding, early exposure to antibiotics, and the timing and type of complementary feeding are the dominant factors (
Figure 2) that influence the composition and function of the gut microbiome in this period and, therefore, determine the development of oral tolerance to different antigens [
48,
49,
50,
51].
Figure 2. Factors shaping the human microbiome development in early life.
The gut microbiome of infants born by cesarean section differs significantly from those born vaginally and thus exposed to their mother’s vaginal and fecal microbes. It is characterized by an enrichment of skin microbes such as
Staphylococcus and
Streptococcus [
52], and a lower microbial diversity dominated by opportunistic pathogens such as
Enterococcus,
Enterobacter, and
Klebsiella species, which are commonly prevalent in hospital settings [
53]. The commensal bacteria that are typically detected in vaginally born infants (i.e.,
Escherichia coli, Bifidobacterium, and
Bacteroides) are significantly lower, and colonization of the intestine by Bacteroidetes is delayed in cesarean-born infants [
52,
53]. These microbiome alterations have been associated with increased susceptibility to specific pathogens and atopic diseases [
54,
55,
56].
Significant differences are also observed in the gut microbiome of infants according to gestational age. The microbiome of preterm infants is characterized by lower microbial diversity and a reduced abundance of strict anaerobes, particularly
Bifidobacterium, compared to facultative anaerobes such as
Enterococcus,
Streptococcus,
Staphylococcus, and various enterobacteria during the first three months of life. [
57,
58,
59]. These alterations are related to several factors, including the immaturity of the immune response, the lack or delay of enteral feeding, and repeated and often prolonged exposure to broad-spectrum antibiotics, which disrupt the gut colonization process in early life and alter the integrity of the mucosal barrier [
60]. As a result, microbial translocation and permeability to microbial components are increased, leading to a higher risk of life-threatening outcomes such as necrotizing enterocolitis and sepsis [
60,
61]. It is noted that gut microbiome diversity in preterm infants decreases with lower gestational age as well as with prolonged periods of antibiotic treatment and parenteral nutrition [
59].
Breast milk is considered the optimal diet for infants and has a beneficial effect on the development of their gut microbiome. In addition to essential macro and micronutrients, it provides prebiotics that promote gut colonization with commensal bacteria and several bioactive factors that modulate host immune responses. As a result, breastfeeding is associated with lower incidence of allergic and autoimmune diseases [
62]. Significant differences are observed in the gut microbiome of infants depending on the type of feeding. The microbiome of breastfed infants is dominated by species of
Bifidobacterium and
Lactobacillus which are known to have immunomodulatory properties and protect against allergies. In contrast, the microbiome of formula-fed infants contains a lower abundance of these beneficial bacteria and is dominated by the
Clostridium,
Granulicatella,
Citrobacter,
Enterobacter, and
Bilophila species [
42,
63]. These differences persist in infants who are still breastfeeding at the age of 12 months and make their “microbiome age” appear younger than that of infants who no longer breastfeed. Therefore, sustained breastfeeding has a strong effect on the formation and succession of gut microbial communities during the first year of life and delays the development of an adult-like gut microbiome [
42,
63]. CMA, particularly allergic proctocolitis, may sometimes occur in breastfed infants. Although there will probably be differences in the gut microbiome between breastfed and formula-fed infants with CMA, there are currently no studies exploring this question.
Increasing evidence suggests that antibiotic administration disrupts the composition of the gut microbiome and impairs its function [
64]. Early antibiotic exposure in infants is associated with reduced diversity and altered composition characterized by a depletion of
Bifidobacterium and a marked increase in the Proteobacteria phylum [
65,
66]. Although weaker, similar alterations have also been found in the microbiome of infants whose mothers received antibiotics before delivery [
53,
65]. Of note, the effect of caesarean section on the infant’s gut microbiome is quite similar to that of maternal antibiotic use during pregnancy [
53].
The Introduction of complementary foods into the diet of infants results in significant changes in their gut microbiome. Solid foods gradually lead to the establishment of an adult-type microbiome dominated by the phyla Bacteroidetes and Firmicutes, represented mainly by the genera
Bacteroides,
Faecalibacterium,
Clostridium, and
Ruminococcus [
42,
48]. In addition to compositional changes, the introduction of complementary foods is also accompanied by functional changes in the microbiome. In particular, it induces a sustained increase in the production of short-chain fatty acids (SCFAs) and promotes the enrichment of the microbiome in genes encoding polysaccharide breakdown, vitamin biosynthesis, and xenobiotic degradation [
67].
In conclusion, studies have shown that the most “beneficial” gut microbiome in infancy is that of exclusively breastfed, vaginally delivered term infants which are characterized by the highest abundance of
Bifidobacterium and the lowest amounts of
Clostridium difficile and
Escherichia coli [
68].
4. The Role of the Gut Microbiome in the Development of FA
Recent data indicate that the composition and metabolic activities of the gut microbiome are inextricably linked to the development of oral tolerance, which is defined as an active process of local and systemic immune unresponsiveness to food antigens [
69]. Studies in animal models revealed that germ-free mice were more likely to be sensitized to CM proteins than those whose gut was colonized with microbes, suggesting a protective effect exerted by the gut microbiome [
70,
71]. Remarkably, the transfer of fecal samples from healthy infants to germ-free mice reduced the risk of CM sensitization and the occurrence of anaphylaxis when exposed to beta-lactalbumin [
72]. These findings suggest that the commensal microbial community drives the host towards the development of oral tolerance. However, the exact mechanism by which this is achieved remains to be further elucidated.
According to accepted theory, the process of oral tolerance occurs in the gut-associated lymphoid tissue (GALT) and involves the recognition of dietary antigens by dendritic cells and the induction of regulatory T cells (Tregs), as well as regulatory B cells (Bregs). Food antigens that cross the epithelial barrier are presented to naïve T cells of the mesenteric lymph nodes by specialized populations of dendritic cells in the presence of retinoic acid and TGF-β. This leads to the robust induction of antigen-specific FOXP3
+ Tregs, which suppress the activation and differentiation of naïve T cells to Th types and have various anti-inflammatory roles. Suppression of Th2 responses also involves Bregs that contribute to oral tolerance by producing specific IgG4 [
73].
Experimental research has shown that the interplay between the gut microbiome and the immune system is crucial for the production of Tregs which are necessary for the establishment and maintenance of oral tolerance. The colonization of the intestine of mice with specific bacterial strains, such as the
Bifidobacterium and
Clostridium species, was shown to have a protective effect on food allergen sensitization by induction of mucosal Tregs [
74,
75]. There are also studies denoting that commensal bacteria, particularly the Clostridia class, are likely to suppress FA through bacterial fermentation products, such as SCFAs, that promote the regulatory activity of dendritic cells and result in the induction of Tregs [
76,
77,
78].
In addition to the induction of antigen specific Tregs, there is evidence that commensal microbes accelerate the development of oral tolerance through their protective effect on the intestinal epithelial barrier [
79]. Indeed, some
Clostridium species have been found to stimulate innate lymphoid cells to produce IL-22, which enhances epithelial barrier integrity and reduces intestinal permeability to protein antigens [
80].
Another factor that has emerged as a crucial determinant for the protection and homeostatic regulation of the intestinal mucosal epithelium is the production of secretory IgA (sIgA), which is the major immunoglobulin on mucosal surfaces. Indeed, reduced fecal IgA levels and altered gut microbiome composition have been associated with food sensitization and an increased risk of anaphylaxis in mice [
81]. The main function of sIgA, referred to as immune exclusion, is to limit the access of pathogens and food allergens to the mucosal barrier, and thus prevents their spread to the systemic compartment [
82]. In addition, it regulates the balance of gut microbes by favoring the maintenance of beneficial members and the removal of opportunistic pathogens, which in turn prevents the absorption of pathogens and toxins [
83,
84]. There is evidence that mucosal secretion of sIgA is partially controlled by the gut microbiome and even that specific microbial species can induce its production to a different extent [
84]. As a result, the composition of gut microbes strongly influences sIgA production and, consequently, the development of oral tolerance.
These data demonstrate a causal role for the gut microbiome in protecting against allergic responses to food antigens. Consequently, alterations in its composition, collectively referred to as dysbiosis, lead to immune dysfunction and predispose the host to the development of FA.