Your browser does not fully support modern features. Please upgrade for a smoother experience.
Please note this is an old version of this entry, which may differ significantly from the current revision.
Subjects:
Oncology
Anaplastic thyroid carcinoma (ATC) are highly aggressive malignant tumors with poor overall prognosis despite multimodal therapy.
- anaplastic thyroid cell carcinoma
- pembrolizumab
- lenvatinib
1. Introduction
Thyroid cancer includes a broad spectrum of histological tumor types and varies from indolent microscopic disease to highly aggressive dedifferentiated tumors [1]. Anaplastic thyroid carcinoma (ATC) comprises approximatively 1% of them and is characterized by rapid growth with local and distant evolution [2]. In the revised 8th edition of the TNM classification, all ATC patients are classified as stage IV and patients with distant metastasis are classified as stage IV C [3]. For the non-metastatic disease, current first-line treatment recommendations are based on surgery radiotherapy and targeted therapies, depending on the presence of the BRAF V600E mutation [4]. Despite this multimodal approach, the prognosis in patients with ATC is poor, with a median overall survival (OS) of 9.5 months [5]. In the metastatic setting, treatment options are limited to systemic treatments and supportive care. Palliation of locoregional disease may be used occasionally to alleviate compressive symptoms and pain in the short term.
Lenvatinib is an antiangiogenic (VEGFR 1–3/FGFR1–4) and antiproliferative (RET/PDGFR) tyrosine kinase inhibitor (TKI), which is approved for differentiated thyroid carcinoma (DTC) refractory to radioiodine treatment [6]. Progression free survival (PFS) for poorly DTC (PDTC) is only 14.8 months and even shorter in ATC [7]. All patients treated with lenvatinib monotherapy ultimately develop treatment resistance and progression [8].
Pembrolizumab is an immune checkpoint inhibitor targeting programmed cell death protein 1 (PD-1) on immune cells. The response to pembrolizumab immunotherapy is associated with the elevated expression of programmed death-ligand 1 (PD-L1) or high tumor mutational burden (TMB) [9,10,11]. In ATC associated with high PD-L1/TMB, the effect of immune checkpoint inhibitors is still low, with 1.9 months of median progression-free survival and 4.4 months of median overall survival (OS) [12]. Rapid tumor development is incompatible with a low response rate and/or a long time to response [13].
The combination of lenvatinib and pembrolizumab is based on a strong mechanistic rationale with immunomodulatory properties: increased tumor infiltration of effector CD8+ T cells and decreased monocytes and macrophages [14]. Adding lenvatinib to immune checkpoint inhibitors (ICIs) may help to overcome primary and acquired resistance to immunotherapy [15]. This combination shows survival benefits in phase-III studies for advanced endometrial cancer after the failure of platinum-based chemotherapy [16] and advanced renal cell carcinoma [17].
2. Preclinical Rationale of Pembrolizumab in Anaplastic Thyroid Carcinoma
The rationale for immunotherapy in ATC is based on the tumor microenvironment, tumor mutation burden (TMB) and Microsatellite instability.
Immune infiltrate Tumor-Associated Macrophages (TAMs), which can make up as much as 70% of the total tumor mass and may function as an immunosuppressive tumor stroma, are a key component of ATC tumors [19,20].
Elevated TAM levels in ATC tumor samples cause a hot immunological environment in 34% of cases, which is characterized by high expression of various inhibitory immune checkpoint mediators, including programmed death-ligand 1 (PD-L1) [4,19]. In both a phase-I investigation of the immune checkpoint inhibitor spartalizumab [20] and a pre-clinical study [21], 70% of ATC samples had PD-L1 expression. In the five cohort publications analyzing PD-L1 expression in ATC, PD-L1 positivity seemed higher in ATC than in DTC or PDTC [22,23,24,25,26]. In addition, CD274 gene amplification is present in 5.1% of ATCs, which is among the ten highest rates of all cancers [27,28]. PD-L1 is a potential predictive biomarker of immunotherapy response in ATC. Spartalizumab, a PD-1 inhibitor, produced an objective response in a recently released ATC cohort from a phase-I/II trial only in patients with detectable PD-L1 expression [20]. The tumor proportion score (TPS) is a PD-L1 measurement in which only membranous staining of tumor cells is regarded as a significant staining. In a recent multicenter research, TPS was detected at 5% in 73% of ATC samples and at least 1% in all ATC [29]. The TPS is a known pembrolizumab response marker established notably for non-small-cell lung cancer and potentially extended to other tumors such as squamous cell carcinoma of head and neck and ATC [30].
Tumor mutational burden (TMB), defined as the total number of somatic mutations per coding area of a tumor genome, is a measure of all non-synonymous coding mutations in a tumor exome [31]. TMB has been shown to vary widely from patient to patient and across tumor types. In striking contrast to PTC, the cancer genome of ATC displays the consequences of genomic instability and is characterized by a significantly higher mutational burden [32]. Comparisons between ATC TMB and TMB in DTCs and PDTCs were analyzed by Pozdeyev et al. using next-generation sequencing (NGS) gene panels in two cohorts of tumor samples (DTC n = 583 and ATC n = 196). Genetic changes were much more prevalent in ATCs than DTCs [33]. Landa et al. sequenced 117 thyroid cancers using NGS including 84 PDTCs and 33 ATCs. Compared to PDTCs, ATCs showed noticeably more mutations [34]. Similarly, 113 tumor samples from DTC, PDTC, and ATC were included in another cohort for TMB analysis, and the results showed that TMB in ATC was greater than in the other subtypes [35]. Several analyses have shown a correlation between high TMB (measured with various methods and cutoff points across studies) and the clinical benefit of immune checkpoint inhibitors [36]. In patients with non-small-cell lung cancer (NSCLC) treated with pembrolizumab as monotherapy, high tissue TMB were associated with durable clinical benefit and extended progression-free survival [37]. A recent biomarker analysis of multi-tumor KEYNOTE-158 study showed that TMB-high status identifies a subgroup of patients who could have a robust tumor response to pembrolizumab monotherapy [11]. TMB could be a novel and useful predictive biomarker for response to pembrolizumab monotherapy in patients with previously treated recurrent or advanced or metastatic solid tumors such as ATC.
Microsatellite instability is caused by the inactivation of the DNA mismatch repair gene(s) encoding mismatch repair (MMR) enzymes MLH1, MLH3, PMS1, and PMS2. This inactivation is a result of ongoing oxidative stress, which also damages the genome and causes poor DNA repair [38]. A genetic analysis of 779 advanced differentiated and anaplastic thyroid cancers noted the presence of MMR DNA deficiency in up to 46% of thyroid cancers with high mutational burden, especially ATCs [33]. A recent literature review revealed the prevalence of microsatellite instability in 7.4% of ATC, with mutations in the MSH2 gene (33%) being the most frequent, followed by MSH6 (25%) and MLH1 (16.7%) occurring in the following combinations: MLH1-MSH2 (8.3%), MSH2-MSH6 (8.3%), and MLH3-MSH5 (8.3%) [39]. In addition, Yoo et al. analyzed somatic copy-number changes in a thyroid tumor samples and showed that they were more common in ATC than in DTC, suggesting that ATC may have a more unstable genome than DTC [35]. Microsatellite instability due to defects in DNA mismatch repair proteins leads to an increase of mutation burdens in cancer-related genes and the formation of neoantigens, which activate the host’s anti-tumor immune response [40,41,42,43]. In cancers treated with immunological check-point inhibitors, mismatch repair deficit or high microsatellite instability has been shown to be strongly correlated with long-term immunotherapy-related responses and a better prognosis in several clinical trials. Pembrolizumab has currently been approved for metastatic tumors with high microsatellite instability or mismatch repair deficiency including ATC [44].
3. Preclinical Rationale of Lenvatinib in Anaplastic Thyroid Carcinoma
Lenvatinib is an oral multi-kinase inhibitor (VEGFR1–3, PDGFR, FGFR1–4, RET, and c-KIT) [8]. Several genetic alterations have been identified in ATC molecular pathways, involving VEGFR1, VEGFR2, EGFR, PDGFRα, and KIT that lead to tumor aggressiveness and progression [45,46]. Ferrari et al. demonstrated that lenvatinib inhibited primary ATC cell cultures proliferation in vitro, while also increasing apoptosis and inhibiting migration and invasion. The antiproliferative effect of lenvatinib in primary ATC cells was observed in all the samples, independently of the absence or presence of BRAF V600E mutation [47]. These outcomes were in accordance with other preclinical studies and showed lenvatinib could prevent angiogenesis by reducing vascular permeability and suppress tumor development in vivo in ATC cell lines [48,49].
RAS-RAF-MAPK, ERK and PI3K pathways are involved in the carcinogenesis of thyroid cancers [50,51] and mutations in these genes are present in ATC [52]. The proteins ERK and AKT were phosphorylated and activated in ATC, making them potential therapeutic targets. Lenvatinib reduced ERK1/2 and AKT phosphorylation in ATC cells, according to research by Ferrari et al. [47].
Lenvatinib also showed evidence of lowering EGFR phosphorylation and inhibiting cell growth via downregulating cyclin D1 expression [53]. Di Desidero et al. demonstrated that Sunitinib, another tyrosine kinase inhibitor (TKI), inhibits Akt and ERK1/2 phosphorylation and down-regulates cyclin-D1 to exert its anti-activated endothelium and ATC cell activity, both in vitro and in vivo [54]. In addition, 67% of ATCs and 77% of ATCs, respectively, were found to express cyclin D1 in studies by Lee et al. [55] and Wiseman et al. [56].
Although many of the TKIs used to treat thyroid cancers that are resistant to radioiodine have similar properties, such as antiangiogenic TKI action, it has been hypothesized that lenvatininb’s greater clinical response may be due to its capacity to also target FGFR1–4 in these rare thyroid tumors [57]. In comparison to normal thyroid and DTC samples, PDTC and ATC samples had the greatest levels of FGFR1 expression. Immunohistochemistry was used by Yamazaki et colleagues to examine FGFR4 expression in 12 ATC patients, and they hypothesized that FGFR4 expression would predict how well lenvatinib would work [58]. Adam and colleagues discovered considerably greater expression of the FGFR1–4 combo in ATC compared to normal thyroid using RNAscope in-situ hybridization [29].
4. Preclinical Rationale of Pembrolizuman and Lenvatinib Association in Anaplastic Thyroid Carcinoma
VEGF-targeted therapies may also function, in part, by blocking VEGF-mediated immune suppression in addition to their anticancer and antiangiogenic effects [59]. In six distinct malignancies, including thyroid carcinomas, immune checkpoint inhibitors and lenvatinib target genes were significantly upregulated and displayed driving alterations, according to a multi-omics investigation [60]. Pathway-enrichment analysis found target genes were implicated in tumor development, angiogenesis, and immunoregulatory associated pathways. These findings are resumed in Figure 1 [60]. Otherwise, lenvatinib and monoclonal antibodies against PD-1 enhanced the immune response in syngeneic mice models. Kato et al. investigated the immunomodulatory activities of lenvatinib in the tumor microenvironment and its mechanisms of enhanced antitumor activity when combined with PD-1 blockade. The antitumor activity of lenvatinib plus anti-PD-1 was greater than that of either single treatment. Flow cytometric analysis revealed that lenvatinib reduced tumor-associated macrophages (TAMs) and increased the percentage of activated CD8+ T cells secreting interferon-γ+ and granzyme B. Combination treatment further increased the percentage of T cells, especially CD8+ T cells, among CD45+ cells and increased interferon-γ+ and granzyme B CD8+ T cells. Transcriptome analyses of tumors resected from treated mice showed that genes specifically regulated by the combination were significantly enriched for type-I interferon signaling [61]. Lenvatinib’s anticancer and immunomodulatory properties were studied by Kimura et al. with a syngeneic hepatocarcinoma mouse tumor model. They showed that lenvatinib had more antitumor activity than sorafenib in a Hepa1-6 tumor model using immunocompetent mice. In addition, CD8+ T cell depletion significantly reduced the antitumor activity of lenvatinib but not sorafenib, indicating that lenvatinib exhibited immunomodulatory activity, particularly on the CD8+ T cell population, and that this effect contributed to lenvatinib’s potent antitumor activity under the immunocompetent condition [14]. Lenvatinib therapy has also been demonstrated to suppress TH2 and boost TH1 immunological responses, by activating memory T cells [62]. Gunda et al. provided evidence for the justification for combining PD-L1 and lenvatinib in ATC using an ATC immunocompetent mice model [63]: lenvatinib caused a noticeable rise in tumor-associated macrophages and tumor-infiltrating immune cells, as well as a noticeable rise in peripheral and tumoral myeloid derived suppressor cells (PMN-MDSC), which together showed dramatic alterations in the immune microenvironment. They came to the conclusion that lenvatinib exhibited pro- and anti-inflammatory effects on the immune system. By reducing the amount of immunosuppressive PMN-MDSC in an experimental manner, they were able to show better antitumoral activity. Inhibition of the PD-1/PD-L1 axis was also associated with a decrease in some immunosuppressive cell types. Thus, lenvatinib plus immune check point inhibitors act synergistically to block PD1/PDL1 axis and the formation of an immunosuppressive microenvironment [64]. Adam et al. looked at the possible immunostimulatory function of FGFR1–4, another Lenvatinib target, in ATC tumor cells. They discovered that the combination of FGFR1–4 was substantially more expressed in ATC than in normal thyroid, but that leukocytes infiltrating the tumor did not express FGFR1–4 [29]. Taken together, these preclinical studies demonstrating lenvatinib’s immunomodulatory activity provide mechanistic rationale for the study of lenvatinib in combination with an anti-PD-1 agent such as pembrolizumab in ATC.
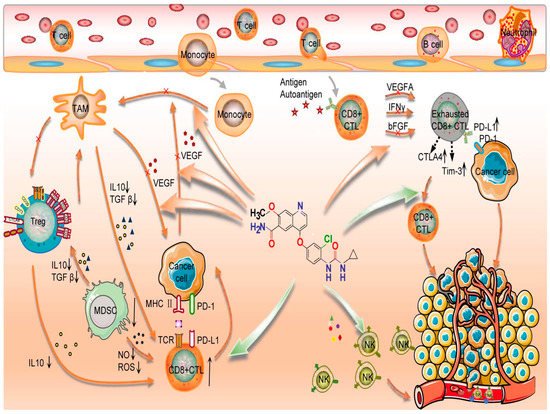
Figure 1. Lenvatinib’s action mode when combined with immunological check point inhibitors. TAM: Tumor associated macrophages; Treg: Regulatory T cells; IL: Interleukine; TGF: transforming growth factor; M DSC: Myeloid-derived suppressor cells; MHC: major histocompatibility complex; IFN: Interferon; ROS: Reactive oxygen species; NO: nitric oxyde; FGF: fibroblast growth factor; CTL: cytotoxic T lymphocyte; VEGF: Vascular Endothelial Growth Factor; NK: Natural killer; TCR: T cel receptor; Tim-3: T-cell immunoglobulin and mucin containing protein-3; PD-1: Programmed cell death protein 1; PD-L1: Programmed death-ligand 1; CTLA 4: Cytotoxique-T-Lymphocyte-Antigen 4 protein; Green arrow: Activation signal; Red arrow: Inhibition signal. Star: Antigen. Cross: Inhibition.
This figure details mechanisms of immunological checkpoint inhibitors combined with lenvatinib. Lenvatinib increased the CD8+T cells function and the cytotoxicity of NK cells, decreased the expression of PD-1, CTLA-4, and TIM3 in T cells, and inhibited T cell exhaustion. Lenvatinib also inhibits tumour angiogenesis and abnormalities by inhibiting the secretion of angiogenic factors, such as VEGF, FGF, and PDGF. Finally, immune checkpoint inhibitors restore the exhausted T cell activity to kill the cancer cell [60].
This entry is adapted from the peer-reviewed paper 10.3390/curroncol29100610
This entry is offline, you can click here to edit this entry!