Your browser does not fully support modern features. Please upgrade for a smoother experience.
Please note this is an old version of this entry, which may differ significantly from the current revision.
Subjects:
Parasitology
|
Immunology
Chagas disease, initiated by the etiological agent Trypanosoma cruzi, is an endemic infection in the American continent. Although vectorial transmission of T. cruzi is recognized as the main mode of infection, other routes such as congenital and blood transfusion are also documented as important methods of transmission. T. cruzi maternal–fetal transmission has been recorded in humans and examined by some investigators in naturally and experimentally infected mammals.
- Trypanosoma cruzi
- Chagas disease
- congenital transmission
1. Introduction
The Trypanosoma cruzi (discovered by Chagas in 1909) (Tc) pathogen is the causative agent of Chagas disease (CD), also known as American Trypanosomiasis (AT) [1]. T. cruzi is a flagellate parasite of the phylum Euglenozoa, class Kinetoplastida, family Trypanosomatidae, and genus Trypanosoma, and it is known to infect humans and various species of wild and domestic mammals [1]. Trypanosomes are unicellular organisms with a single nucleus located in the center of the body. The infectious trypomastigote form of T. cruzi is typically spindle-shaped with an undulating membrane along its axis and a DNA-rich mitochondrial organelle called kinetoplast located in the anterior part of the parasite [2]. A series of microtubules originating near the kinetoplast make up the basal body that extends along the undulating membrane to the opposite end of the parasite. The flagellar tubules are surrounded by a membrane, and together they form the flagellar pocket [3]. Trypanosomes contain many of the same organelles as noted in their mammalian host, but also have organelles, e.g., glycosome, acidocalcisome, cytoplasmic vacuole, and reservosome, that are unique to these parasites [3,4].
Patients exposed to T. cruzi encounter a high parasitic load in circulation and various tissues for 1–2 months, after which it is significantly controlled by the adaptive immune response, and then intermittent low level of parasites are observed during the indeterminate phase of the disease [5]. Progression to clinically symptomatic chronic disease phase in ~30–40% of the infected patients is presented with the development of cardiomyopathy, though other morbidities including gastroenterological and neurological disorders are also noted in Chagas patients. Chagas disease can ultimately cause death due to heart failure [6].
T. cruzi populations survive in a wide ecological range and exhibit a highly diversified genetic profile. By analyzing a variety of molecular markers, T. cruzi clones have been categorized into seven discrete typing units (DTUs) referred as TcI-TcVI and Tcbat [7,8,9,10]. The parasite isolates belonging to these DTUs differ in geographical distribution and epidemiological association, as well as in virulence, pathogenicity, and tissue tropism in the host [11,12,13]. In Mexico and Central America, T. cruzi isolates of TcI DTU are found with the highest prevalence [14], though mixed infections with TcII, TcV, and TcVI are also noted in Mexico and Central America [15]. T. cruzi isolates of TcI and TcII DTUs with some cases of TcIV, TcV, and TcVI DTUs are noted in Brazil [16] and Argentina [11,17]. TcI-TcVI DTUs have been reported in large number of vectors species and mammals, Tcbat is transmitted from infected bats, and all DTUs can cause Chagas disease in humans and animals, though some evidence indicates an association between DTUs and clinical outcomes [18]. Readers interested in more details of geographic distribution of T. cruzi lineages in mammalian and vectorial hosts are referred to recent review articles [9,19].
The World Health Organization estimates that approximately 6–7 million people are affected by Chagas disease worldwide, and nearly 100 million people live in the endemic areas with high risk of exposure to T. cruzi infection in the Latin American countries [20,21]. The main form of transmission is by contact with feces of infected triatomine insects. Infection occurs when T. cruzi-infected triatomines feed on the host, contaminated feces from the insect is excreted, and parasites gain access to the blood stream of the host via skin puncture or mucous membranes [22]. Parasite transmission may also occur by oral ingestion of infected bugs [23], transfusion of infected blood [24], and congenital transmission from infected mothers [25]. After the vector control programs implemented in 1980–1990s were highly successful in controlling the acute transmission of T. cruzi from infected bugs in South America, the non-vectorial transmission pathways have emerged as major issues of public health concern [24]. In fact, congenital transmission has led to the globalization of Chagas disease in non-endemic countries [26,27,28,29]. It is projected that internationally over two million women of reproductive age are infected with T. cruzi, and 1–10% of fetuses carried by infected mothers are born with CD [30,31,32].
In this review, we summarized the current knowledge regarding T. cruzi congenital transmission in humans and animal models with an aim to point out the similarities and dissimilarities of placental and maternal factors between animals and humans. It is our hope that the current review will serve as an essential source of knowledge for understanding the pathophysiology of congenital transmission and permit the reader to make an informed decision when using experimental models of T. cruzi infection for studying the pathomechanisms of congenital transmission or examining the efficacy of new diagnostic tests or therapies to prevent congenital CD.
2. Diagnosis and Burden of Congenital Transmission of T. cruzi in Humans
Women, irrespective of being in the acute or chronic phases of CD, can congenitally transmit the parasite to the fetus and newborn [33]. Yet, there is significant evidence that a high rate of transmission occurs when pregnant women are acutely infected and/or develop reactivated acute infection due to immunosuppression [34]. Indeed, ~53% of women who became infected during pregnancy exhibited prenatal or perinatal transmission of T. cruzi to their fetuses and newborns [34,35,36,37]. Likewise, women infected with T. cruzi prior to pregnancy who became exposed to HIV or were treated with immunosuppressive drugs during gestation transmitted T. cruzi with high frequency to their newborns [38,39]. Besides, high parasite burden in umbilical cord of infants is associated with the severity of congenital CD [31,40]. In Mexico, studies indicate that an estimated seroprevalence of 2.21% (95% CI 1.46–2.96) would result in 50,675 births from T. cruzi-infected pregnant women and ~3193 cases of vertical transmission and infected neonates per year [41]. It is of note that infected mothers transmitted the same DTUs of T. cruzi to their newborns as were predominantly identified in the local population [17,42,43]. When pregnant women were exposed to mixed or multiclonal infections, predominance of different clones in the mother and their newborns has been noted [42,44,45]. Overall, the current literature allows us to surmise that (a) all DTUs can potentially be transmitted via congenital route, (b) natural selection of the transmitted parasite may occur during pregnancy, and (c) diagnostic screening of pregnant women and newborns and identification of parasite lineage would inform the timely treatment of newborns and restrict T. cruzi transmission.
The criteria for the occurrence of congenital transmission in humans are that the mother is T. cruzi-seropositive, parasites are detectable in the peripheral blood of the newborn, and anti-T. cruzi antibodies are made in the newborn after passive immunity acquired during lactation has disappeared (if vector and blood transfusion infection have also been ruled out) [37,46]. Microscopic observation of parasite in fresh blood smears offers the simplest approach to diagnosis of T. cruzi infection in newborns [47]. When there is a low number of blood parasites, concentration methods such as Strout or micro Strout test are useful [48,49]. Another form of T. cruzi detection is micro hematocrit, which may be more convenient for parasite detection because of its simplicity, low cost, and capacity to detect 40 parasites/mL with 97.4% sensitivity [50]. However, parasite identification with micro hematocrit requires ~30 min per sample evaluation by trained personnel, and even then, this method identifies only 40–60% of the congenital transmission in newborns [46,51]. Indirect parasite detection methods such as hemoculture or xenodiagnosis, although sensitive, have the disadvantage in that they require several weeks for positive T. cruzi identification [48,52].
In infants of nine months age or older, serological detection of anti-T. cruzi antibodies by indirect hemagglutination assay (IHA) [53], indirect immunofluorescence (IIF) assay [54,55], or enzyme-linked immunosorbent assay (ELISA) [52,53] can be employed. Serology is routinely applied for diagnosis of T. cruzi infection in clinical laboratories in Latin America [48,52,56]; however, difficulty in following up the newborns is a major limitation of this method in diagnosing and treating the infected infants in a timely manner.
The application of conventional polymerase chain reaction (PCR) for positive or negative diagnosis of congenital T. cruzi infection has increased in recent years [48,56,57]. When used at birth, PCR test provides higher sensitivity and specificity compared with other parasitological identification methods [37,58]. Indeed, accuracy of PCR test in diagnosis of congenital T. cruzi infection has been demonstrated in several studies [56,59,60,61]. A variation of PCR is the quantitative PCR (qPCR), which allows enumerating the parasite burden [62]. The qPCR assay based on T. cruzi satellite DNA and kinetoplast DNA can detect 0.85 and 0.43 parasite equivalents per mL blood, respectively, making it a highly sensitive approach [63]. The main drawback of this technique is the requirement for specific laboratory instrument and well-qualified employees who maintain rigorous quality control. As an alternative, loop-mediated isothermal amplification (LAMP) can be performed using a heat-block at a constant temperature of 60–65 °C and it does not require specialized PCR equipment [64,65]. LAMP has been successfully implemented to amplify T. cruzi DNA with a similar sensitivity as was noted with qPCR [64,66]. One caveat in implementation of molecular assays in the field is that trained personnel are needed to accurately identify infected newborns and avoid false-positives due to contamination of maternal parasite DNA. Readers are directed to a recent review discussing the efficacy and implementation of old and new diagnostic tests and an ideal algorithm for diagnosis of congenital transmission of T. cruzi in infants [25].
3. Characteristics and Classification of the Mammalian Placental Barrier
The placenta is a temporary organ formed during pregnancy. It primarily functions in anchoring the fetus to the uterine wall and mediating the immune tolerance to avoid immunological rejection of the fetus while also maintaining the anti-infectious capacity [67,68,69]. Placenta is also required for the transfer of nutrients such as amino acids, lipids, and glucose to the fetus, and the exchange of oxygen, carbon dioxide, and fetal waste excretion [70].
The two forms of placenta classification are based on gross shape and histological structure (Figure 1). Gross morphology based placental classification describes whether maternal–fetal exchange occurs on all of the available surface of the chorionic sac or if it is restricted to specific zones [71,72]. Accordingly, four types of placentas categorized based on gross shape are shown in Figure 1A. These include (1) diffuse placenta, which appears over the complete surface of the uterine luminal epithelium with the formation of folds and villi, and it is seen in horses and pigs; (2) cotyledonary placenta, which consists of numerous spot-like placental regions of the endometrium known as caruncles or cotyledons with smooth and avascular intervening areas in the chorion, and it is seen in ruminants; (3) zonary placenta, which shows an intimate interdigitating contact zone that forms a strap or girdle around the chorionic sac, and it is seen in carnivores such as dogs, cats, bats, seals and bears; and (4) discoid/bidiscoid placenta, which may contain a single (discoid) or double (bidiscoid) disc, wherein maternal–placental interaction occurs in a roughly circular area, and it is seen in humans, rodents, rabbits, and primates [71,73].
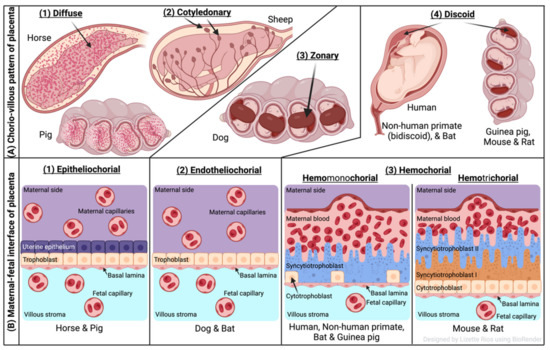
Figure 1. Comparative features of mammalian placenta. (A) Chorio-villous patterns of placenta. There are four types of placentas that are recognized: (1) diffuse: occurs over the entire surface of the uterine luminal epithelium with formation of folds/villi (seen in horses and pigs); (2) cotyledonary: characterized by many spot-like placental regions of the endometrium known as caruncles or cotyledons with smooth and poor vascularized intervening areas in the chorion (seen in ruminants); (3) zonary: placenta shows an intimate interdigitating contact zone that forms a belt or girdle around the chorionic sac (seen in carnivores such as dogs); (4) discoid/bidiscoid: characterized by a single (discoid) or double disc (bidiscoid), and mother–product interaction is confined to a roughly circular area (seen in humans, rodents, bats, and primates). (B) Maternal–fetal interface of placenta. This classification of placentas depends on the cell layers between the tissues of the mother and the fetus. (1) Epitheliochorial placenta (seen in horse, pig, and ruminants) is the least intimate because the interaction between the maternal blood and the fetal tissue is limited by a layer of uterine epithelial cells and a layer of trophoblast cells. (2) Endotheliochorial type (seen in dogs and cats) is the second more invasive barrier, where the uterine epithelium in degraded after implantation, leaving to the trophoblast adjacent only to the maternal endothelium. (3) Hemochorial barrier (hemomonochorial in primates and hemotrichorial in rodents) is the most intimate interface. The hemochorial epithelial and endothelial cells of the mother are degraded, leaving the trophoblast cells (syncytiotrophoblast and cytotrophoblast) in direct contact with the maternal blood.
Three main types of placental classification were proposed by Grosser based on histological structure (Figure 1B) [74]. This classification recognizes the histologic relationship of the chorion and uterine wall, and therefore, it is more apt and informative in describing the placental function [71,72]. The epitheliochorial type of placenta (seen in horses, pigs, and ruminants) is most superficial or least intimate because the maternal blood—fetal tissue interactions are limited by layers of uterine epithelial cells and fetal trophoblast cells only. In endotheliochorial placenta, withdrawal of maternal uterine epithelium and connective tissue after implantation informs the maternal endometrial contact with fetal trophoblasts. The endotheliochorial type of placenta is seen in four major clades of eutherian mammals, including carnivores, dogs, and cats. Lastly, the hemochorial placenta is considered most invasive, as in this case maternal hemochorial epithelial and endothelial cells are degraded, and thereby maternal blood is in open exchange with fetal trophoblast cells (syncytiotrophoblast and cytrotrophoblast). The hemomonochorial, hemodichorial, and hemotrichorial placentas consist of one, two, and three trophoblast layers, and are noted in primates, rabbits, and rodents, respectively [73,75].
This entry is adapted from the peer-reviewed paper 10.3390/pathogens11101172
This entry is offline, you can click here to edit this entry!