Meiosis is the unique division of germ cells resulting in the recombination of the maternal and paternal genomes and the production of haploid gametes. In mammals, it begins during the fetal life in females and during puberty in males. In both cases, entering meiosis requires a timely switch from the mitotic to the meiotic cell cycle and the transition from a potential pluripotent status to meiotic differentiation. Revealing the molecular mechanisms underlying these interrelated processes represents the essence in understanding the beginning of meiosis. Meiosis facilitates diversity across individuals and acts as a fundamental driver of evolution. Major differences between sexes and among species complicate the understanding of how meiosis begins.
1. Introduction
Meiosis is a special type of cell division unique of the germ cells of sexually reproducing species implying paternal and maternal genomic recombination and resulting in the production of haploid gametes that at fertilization fuse to reestablish normal diploidy in the zygote. It involves one round of DNA replication followed by two rounds of cell divisions, resulting in the mammalian male in four haploid germ cells, termed spermatids, and in the female in one mature oocyte and three small cells, termed polar bodies. Meiosis comprises two stages, based on the two rounds of divisions: meiosis I and meiosis II. Each stage can be subdivided into prophase, metaphase, anaphase, and telophase. Prophase I comprises events exclusive for meiosis and can be subdivided into four stages: leptotene (chromosome condensation), zygotene (the pairing of homologous chromosomes through the synaptonemal complex), pachytene (the initiation of the recombination between chromosome pairs and the formation of chiasmata between homologues at the crossing-over sites), and diplotene (chromosomes begin to separate, but remain attached via chiasmata).
2. Molecules and Conditions for Meiotic Beginning
2.1. Intrinsic Factors
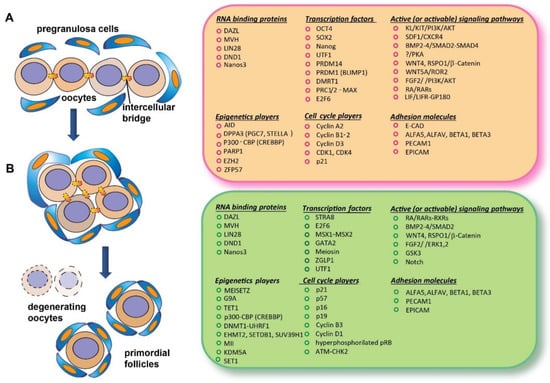
The beginning of meiosis in germ cells requires the acquisition of a molecular status, critically including epigenomic and specific RNA-binding proteins, referable to what early researcher conceived as “intrinsic factors” (features not directly traceable to the action of specific external signaling) and a variety of local signals and conditions constituting the “extrinsic factors” (Figure 1).
Figure 1. Schematic representation of an oogonium nest before entering meiosis (A) and of oocytes at the beginning of meiosis associated with nest breakdown and the formation of primordial follicles (B); on the right, the main classes of molecules identified in mouse oocytes during these developmental stages.
2.1.1. Genome-Wide Reprogramming in Premeiotic PGCs
Early studies revealed that, in both sexes, the DNA of mouse PGCs settled in GRs was strikingly hypomethylated and that this epigenetic erasure involved imprinted genes [
5,
38]. Today, we know that this unique epigenetic status is part of a global genome-wide reprogramming proper of the preceding germline and probably necessary for the beginning of meiosis.
PGC genome reprogramming relies on epigenetic modification, including global and locus-specific DNA demethylation and histone replacement, and post-translation modifications.
To these epigenetic processes, the recruitment of inhibitory and activatory complexes on specific gene loci are superimposed. The biological outcomes of these processes are multiple. They prevent the inheritance of acquired epigenetic information across generations and erase parental imprints so that they can be re-imposed in a sex-dependent manner. In addition, they reactivate loci on the inactive X in female germ cells and probably establish histone marks, making it possible to rapidly regain totipotency in the zygote and blastomeres.
In mouse PGCs, global and locus-specific DNA demethylation occurs in two subsequent phases [
40]. The first stage happens in specified PGCs during their proliferation and migration towards GRs (7.5–10.5 dpc) [
41,
42].
Active and passive mechanisms of DNA demethylation work in concert. The first is likely mediated by Activation-induced cytidine deaminase (AID) and perhaps by Poly-(ADP-ribose) polymerase 1 (PARP1) [
43,
44,
45]. The second one is related to DNA replication occurring during PGC proliferation in the absence of key components of the methylation machinery [
41,
46]. In this phase, transcription factors such PRDM1 (or BLIMP1) and PRDM14 (SOX17 in humans) [
47] repress somatic transcriptional programs, while genes encoding core pluripotency transcription factors such as OCT4, SOX2, and Nanog, and germ-cell-specific genes (i.e.,
Nanos3) are activated [
48].
Notably, during this phase, methyltransferase DNMT1 preserves DNA methylation in the differentially methylated regions (DMRs) of some imprinted genes, cytosine guanine island (CGI) promoters on the inactive female X chromosome, and of germ-cell-specific and meiotic genes [
49]. Repressive histone marks H3K9me2, H3K27me3, and H2A/H4R3me2s, together with H2A.Z, and active histone marks H3K9ac, H3K4me2, and H3K4me3 contribute to refining the chromatin status of pre-gonadal PGCs [
5,
50,
51,
52]. In addition, catalytically active Polycomb repressive complex ½ (PRC1/2) was identified in 12.5 dpc PGCs. These chromatin remodelers are required to repress meiotic genes,
Stra8 included, from depositing repressive histone marks on their promoters [
53].
The second reprogramming occurs after PGCs enter the GRs (11.5–13.5 dpc). The loss of repressive histone marks likely favors an open chromatin status [
5]. As noted, among the repressive histones, the decrease in H3K27me3 is followed by an increase from 12.5 to 16.5 dpc concomitant with the beginning of meiosis in female germ cells [
54], suggesting that a high level of this histone may be necessary for meiotic entry. Notably, histone modifications seem to be, at least in part, sex specific [
55]. Interestingly, mouse PGCs obtained from 11.5–13.5 dpc specimens of both sexes revealed H3K4me3/H3K27me3 bivalent domains to be highly enriched at the developmental regulatory genes in a manner remarkably similar to that of embryonic stem cells (ESCs) [
56]. Bivalent modification can contribute to the repression of somatic cell lineage genes and/or ready these genes for future activation.
Notably, locus-specific DNA demethylations, including germ-cell-specific and meiotic genes, occur in PGCs of both sexes so that most of the sequences previously maintained methylated by DNMT1 are demethylated [
5,
41,
57]. Critically, this induces the expression of
Dazl and others germ-cell-specific genes, such as
Mvh and
Sycp3 [
57,
58].
TET1/2, the enzyme capable of converting 5-methylcytosine (5mC) into 5-hydroxy-methylcytosine (5hmC) [
59], plays a critical role in these second demethylation round [
57,
60,
61], perhaps with the participation of PARP1 [
44]. TET1 seems, however, mostly devoted to the removal of aberrant residual and/or de novo DNA methylation and to the activation of germ-cell-specific and meiotic genes via a DNA demethylation-independent mechanism [
61,
62]. Interestingly, DAZL, which is essential for licensing gametogenesis (see below), associates with the mRNA of
Tet1 in mouse ESCs supporting its translation [
63].
Collectively, these data suggest that one purpose of the epigenetic reprogramming in PGCs, besides those reported at the beginning of this section, is to ensure the timely and efficient activation of genes enabling progression towards gametogenesis and meiosis.
In such a view, epigenetic modifications might represent, at least in part, the hypothesized intrinsic factors governing the beginning of meiosis in female germ cells (see above). In supporting this notion, the ATP-dependent SWI/SNF remodeling chromatin complex counteracting PRC-mediated repression is required for the transition from sexually undifferentiated mouse PGCs to female or male germ cells [
64]. In addition, Hill et al. identified a subset of genes activated during PGC epigenetic reprogramming, which are referred to as the Germline Reprogramming Responsive (GRR) genes that are activated after the depletion of 5mC and PRC1 in both male and female PGCs at E13.5 [
62].
In the mouse embryo, following initial inductive triggering by growth factors such BMPs and WNT3a needed for germline specification inside epiblast cells [
65,
66,
67,
68,
69], epigenetic reprogramming proceeds in the PGC precursors and subsequently in PGCs, provided that growth factors/cytokines sustain their proliferation and survival during migration and the colonization of GRs [
70,
71]. Understanding if and how signaling from the above pathways and environmental cues modulate the chromatin-modifying enzymes represents a challenge for future research.
2.1.2. DAZL and MVH
Substantial evidence supports the notion that in mice and probably in all mammalian species, RBP DAZL is critical for rendering female PGCs/oogonia competent to enter meiosis within the fetal ovary and for sex-specific differentiation events of oogenesis and spermatogenesis [
80,
81,
82]. Despite this, the gametogenic functions of DAZL have not yet been fully characterized. DAZL was localized in both the nucleus and cytoplasm of fetal germ cells [
83]. As reported in the previous section, its expression is epigenetically regulated by the DNA methylation of CpG islands present in its promoter region. Moreover, the fact that the first exon and intron of the gene are highly enriched in the active histone H3K4me3 mark and lack repressive H3K27me3 [
84] suggests that promoter demethylation can rapidly lead to gene activation. The expression of the
Dazl gene, however, might also depend on signals from the gonadal ridges, since in the absence of these, PGCs of both sexes were shown not to undergo gametogenesis and to retain their pre-gonadal characteristics [
58].
Dazl and
Mvh (see below) are not likely direct targets of RA, one of the major extrinsic factors triggering meiosis in mammalian germ cells, since they were not reported to possess Retinoic Acid Response Elements (RAREs) in mammals. Intriguingly, however, a functional Retinoic Acid Receptor beta (RARβ)-binding RARE sequence was found in the chicken
Dazl gene core promoter region [
85].
Deleted in azoospermia, the DAZ family consists of three members, DAZL, DAZ, and BOULE, which are exclusively expressed in pluripotent stem cells and germ cells [
88]. As RBPs, these factors elicit their roles through modulating the translation and stabilization of specific mRNAs [
89]. For example, DAZL, either directly, through 3′-UTR, or indirectly, through polyA-binding protein PABP, binds the mRNAs of germ-cell-specific genes, such as
Mvh,
Sycp1,
Sycp3,
Tex11, and
Tex14, and facilitates the translation of their coded proteins [
89,
90].
Notably, in mouse embryos, depending on the genetic background, the ablation of
Dazl causes a severe reduction in germ-cell numbers and the aberrant expression of markers of pluripotency in post-migratory PGCs. Moreover, these cells show the inability to differentiate along the oogenesis or spermatogenesis pathway and in female germ cells to enter or progress through meiosis [
4,
80,
81].
By profiling gene expression in mouse fetal ovary mutants, Soh et al. recently reported that DAZL is required for the induction of nearly all 104 genes that they identified to be specifically expressed during meiotic prophase [
91], but the way in which it controls such transcriptional program remains to be clarified. A study documented that GASZ, a protein with four ankyrin repeats encoded by an evolutionarily conserved gene expressed exclusively in germ cells, interacts with DAZL and synergistically stimulates PGCLCs to form mouse ESCs [
92].
Evidence exists indicating that DAZL also plays a crucial role in human oogenesis [
93]. In the human embryo, the percentage of germ cells in the fetal ovary highly expressing DAZL was found to be increased markedly from 28 to 48% from weeks 10 to 18 of development, while that of OCT4-positive cells was found to be decreased significantly. Notably, cells expressing a high level of DAZL almost always lacked OCT4 expression, indicating a mutually exclusive expression pattern of the pluripotency marker and DAZL [
94].
Moreover, a combination of DAZL and BOULE could be utilized to induce human ESCs to exit the pluripotent state and enter meiosis in vitro [
94]. The identification of RNA codifying genes involved in chromosome cohesion and DNA recombination as targets of human DAZL (
SYCP3 and
TEX19, for example) highlights the importance of this RBP also in early meiosis [
95].
Similar to
Dazl,
Mvh (also known as
Vasa or
Ddx4) is expressed in germ cells around the time of PGC arrival at the gonadal ridges and requires the gonad environment for expression [
96].
Although
Mvh expression is independent of DAZL [
58], the latter is able to bind
Mvh RNA and modulate its translation [
90]. Moreover, as reported above,
Mvh expression is partly dependent on demethylation [
42].
MVH is an ATP-dependent RNA helicase that often changes the secondary structures of RNA during processes such as alternative splicing and protein translation initiation [
97]. MVH interacts with ribonucleic acids through its conserved DEAD (Asp-Glu-Ala-Asp)-RNA-binding motif and exerts multiple roles in the germline of various species [
98]. For example, it appears to be able to regulate the proliferation and pluripotency of PGCs and meiosis in male germ cells; however, no specific functions of the protein in mammal oogenesis were found. Indeed, the loss of
Mvh in the mouse affects the number and differentiation of male germ cells but apparently not oogenesis [
99].
2.2. Extrinsic Factors
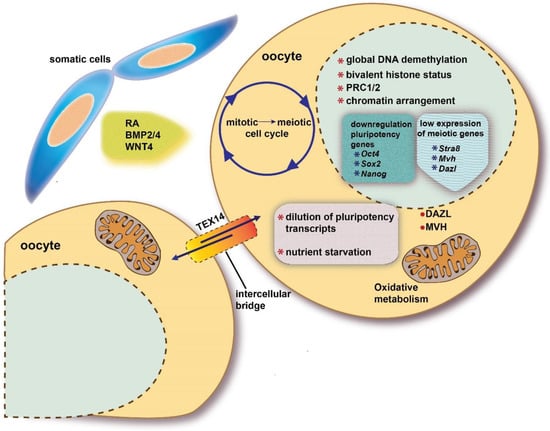
Once a specific epigenetic status and the expression of DAZL RBP are established, PGCs become responsive to the action of local ovary/mesonephros factors and conditions that though various intracellular molecular cascades, promote or inhibit in the developing ovaries and testes, respectively, their entering into meiotic prophase I. Such extrinsic factors cause a mitotic G0 block in male PGCs/prospermatogonia, and a switch from the mitotic to meiotic cycle and the full activation of meiotic genes in female PGCs/oogonia (Figure 2).
Figure 2. Schematic representation of intrinsic and extrinsic factors discussed in the present review involved in the beginning of meiosis in a fetal oocyte.
2.2.1. The Switch from Mitotic to Meiotic Cell Cycle
The in vitro transdifferentiation of isolated premeiotic PGCs into pluripotent stem cells, termed EG cells, by specific cocktails of growth factors (KL, FGF2, LIF) or GSK3 and MAPK inhibitors (2i) [
100] and the formation of Embryonal Carcinoma (EC) cells in vivo from undifferentiated PGCs [
101] confirm the essential influence of signals from the fetal gonad environment on directing the correct germ-cell differentiation and meiosis. Moreover, these occurrences offer some clues on the processes that critically control the timely entering of PGCs into meiosis in fetal ovaries and into mitotic arrest in fetal testes. Among these there are the already mentioned epigenetic genome status and the downregulation of pluripotency genes associated with the activity of intracellular pathways depending on external signaling demonstrated to drive, at least in vitro, PGC proliferation/survival. These include KL/KIT-dependent PI3K/AKT pathways, PKA and ERK1/2 pathways, and PKCε, a novel DAG-dependent PKC. The manipulation of PGC epigenetics (i.e., the inhibition of deacetylation) or of signal transduction pathway components (i.e., the constitutive expression of AKT and the addition to the culture medium of FGF2, FRSK, PKCε inhibitor, or 2i) prevents the beginning of meiosis and makes both female and male PGCs prone to transdifferentiate in proliferating EG cells [
100]. Interestingly, most of these molecular pathways seem to be present and active in human PGCs during the transition to premeiotic oogonia [
102].
Regarding the G1 phase, early studies by Western et al., 2008, showed that in female mouse premeiotic PGCs, the critical regulator of mitotic G1–S transition, pRB, was present but became hyperphosphorylated and inactive, while another member of the RB family,
Rbl1, was downregulated [
103]. More recent studies in mouse and human germ cells revealed that the transcripts of other players in the mitotic G1–S transition, such as
Ccnd1, were low in mitotic germ cells and sharply increased in early meiotic cells, whereas
Ccna1 was exclusively present in meiotic germ cells [
104]. The expression of
Ccna2 and
Ccnd3, both also exerting a role in the G2–M transition, was restricted to mitotic germ cells and downregulated in meiotic prophase.
Cdk1 and
Cdk4 transcripts appeared to be restricted to mitosis, while
Cdkn2a (p16) and
Cdkn2d (p19), inhibitors of cyclin D/Cdk4/Cdk6 complexes, increased consistently with
Ccnd1 in early meiotic germ cells.
Conversely, Cdkn1a (p21) and Cdkn1c (p57), inhibitors of most cyclin/Cdk complexes, decreased during the transition from mitosis to meiosis. Whether some of these changes contribute to meiosis entry or are a consequence of the change in the cell division process remain to be established.
In the G1 phase of the cell cycle, many DNA replication regulatory processes begin. In the case of meiosis, DNA replication during the S phase produces pairs of sister chromatids, held together by cohesin complexes. Much of our understanding of this pre-meiotic DNA replication comes from studies using yeasts, which reported several differences between mitotic DNA replication and pre-meiotic DNA [
105,
106,
107]. The pre-meiotic S phase is also of longer duration than the mitotic S phase in mammals (from two to three times) [
108,
109,
110]. This substantial extension of the pre-meiotic S-phase is still largely unexplained, although the synthesis of proteins such as STRA8, DMC1, SYCP3, Hormad1, and REC8, needed for setting up the inter-homologous relationships important for the recombination and correct segregation of homologous chromosomes, is a possible reason. Recently, STRA8, as well as its germ-cell-specific interactor, Meiosin, emerged as the protein required for the proper expression of many meiotic cell-cycle genes [
111,
112]. We discuss the involvement of STRA8 and Meiosin in the mitosis/meiotic switching operating in this phase in another section below.
2.2.2. Retinoic Acid
As anticipated in the previous section, multiple signals from the somatic cells of the gonad–mesonephros region are necessary for correctly promoting meiosis. Despite some criticisms (see above and [
32,
116,
117,
118]), several in vivo and in vitro studies indicated that RA is involved in triggering or permitting meiosis in a paracrine manner in mammal germ cells of both sexes [
119].
In the signaling cells, the conversion of retinol to RA requires two sequential oxidative steps, catalyzed by retinol or alcohol dehydrogenases (RDHs or ADHs) and by retinaldehyde dehydrogenases (RALDHs), respectively. Cytochrome P450 enzymes CYP26A1, CYP26B1, and CYP26C1 finely control the level of RA present in each tissue, balancing its synthesis and degradation [
120].
In mice, a flux of RA responsible for the beginning of meiosis in the fetal ovaries is believed to come mainly from the mesonephros between 11.5 and 14.5 dpc [
7]. A local ovary contribution to RA production is also possible [
128,
129]. A rostro-caudal wave of expression of genes such as
Dazl,
Stra8,
Sycp3, and
Rec8 is typically described in mouse female germ cells between 13.5 and 15.5 dpc, whereas that of pluripotency markers is quenched in the same direction [
1,
2]. In fact, the expression of a large number of genes is modulated in the transit of PGCs to primary oocytes in the mouse ovary between 12.5 and 15.5 dpc. As reported above, this includes pluripotency and meiotic prophase genes, and RA likely plays a crucial role in modulating both classes of genes.
Of note, this transcriptional program characterizing MPI beginning in the ovary is very similar to that engaged by preleptotene spermatocytes [
111,
112], and it was highlighted that the expression of more than 50% of them is STRA8 dependent [
42,
91,
112].
In both mice and humans,
Stra8 is considered a major RA target in germ cells entering meiosis. How RA induces
Stra8 expression is, however, still the object of debate. As reported above, in the classical model of RA-dependent gene activation, unliganded RAR–RXR heterodimers bind to RARE sequences and repress the transcription of their associated genes, unless activated by RA binding. Additional co-regulators and epigenetic changes critically contribute to transcriptional regulation mediated by RA. All such components are probably involved in the RA-dependent stimulation of
Stra8 expression. Studies in mice indicated that
Stra8 possesses classical RARE sequences, but its involvement in
Stra8 activation by RA is not completely clear [
133,
134].
2.2.3. STRA8
Stra8 was first identified in P19 teratocarcinoma cells as a RA-inducible gene and erroneously described as being restricted to male germ cells in fetal testes [
143].
Gene knockout studies in mice clearly demonstrated that
Stra8 is required for meiotic initiation and the meiotic progression of germ cells and that the ablation of the gene results in infertility in both sexes [
6,
7,
8,
9,
10]. In mouse female embryos,
Stra8-deficient PGCs do not initiate meiotic chromosome condensation, cohesion, and synapsis, or DNA double-strand breaks and recombination [
6].
Additionally, different transcriptional (e.g.,
Pparg) and post-transcriptional regulators (e.g.,
Meioc and
Ythdc2) involved in the control of meiotic prophase length (see above) are STRA8-regulated [
91,
111,
112], thus indicating that other factors could participate in the STRA8 regulation of the meiotic program. In fact, how STRA8 exercises its transcriptional control is not completely clear. It is able to directly bind the genomic regulatory region close to the transcriptional start site (TSS) of the regulated genes [
111,
112] at a consensus motif (the CNCCTCAG sequence) that does not correspond to a E-Box sequence recognized by bHLH transcription regulators [
155].
The same consensus region in the most meiotic-regulated genes is shared by Meiosin, a recently discovered STRA8 interactor in prospermatocytes [
111]. This protein is also expressed in the mouse fetal ovary at the same time as STRA8, and ovarian germ cells fail to start meiosis when
Meiosin is ablated as in
Meiosin-/- spermatocytes. In addition, Meiosin, which possesses bHLH and HMG DNA-binding domains, similar to STRA8, controls a large transcriptional program, including meiotic and non-meiotic genes, most of which are also STRA8-regulated, thus indicating that Meiosin and STRA8 act in concert to amplify the network, ensuring the exact gene level indispensable for the beginning of meiosis at the right time. Do they conduct this by binding to open chromatin and recruiting RNA polymerase II, releasing it, or via other mechanisms? This is still an open question.
2.3. Other Factors
2.3.1. Inhibitory and Cooperative Factors
The downregulation of the FGF9 and NODAL growth factors is considered as a prerequisite to make meiosis possible in the mouse embryonic ovary. FGF9, expressed in the somatic cells of both male and female undifferentiated GRs, remains highly expressed only in fetal testes, specifically in pre-Sertoli cells, mainly exerting a paracrine action on germ cells [
29,
159]. Conversely, NODAL, a member of the TGFβ superfamily, appears to be mainly expressed in PGCs and might act in an autocrine manner [
137,
160].
Intriguingly, in the human embryo, NODAL was found to be expressed in both female and male mitotic germ cells, whereas its receptor, ACVR1C, and target gene
Pitx2 were specifically expressed in female meiotic and male mitotic-arrest germ cells [
161]. Nonetheless, at least in the mouse, the absence of either FGF9 or NODAL can be considered permissive for meiosis onset rather than inductive.
β-Catenin was also recently proposed to exert a negative action on the beginning of meiosis. Indeed, Le Rolle et al. [
162] reported that β-Catenin was present in the nucleus of proliferating mouse PGCs physically associated with OCT4 and regulated chromatin accessibility to promoters and intergenic regions of several genes. The ablation of
β-Catenin caused the exit from the mitotic cycle and the precocious expression of
Dazl and meiotic genes including
Stra8. In light of these and other results, the authors proposed that the activation of GSK3 and the upregulation of ZNRF3, an E3 ubiquitin-protein ligase that acts as a negative regulator of the WNT signaling pathway, in PGCs promote the inactivation of β-Catenin and contribute to the switch from mitosis to meiosis.
Conversely, Activin A might cooperate with RA to promote meiosis. When Activin A was added to cultured 12.5 dpc mouse ovaries or delivered to 10.5 dpc litters via intraperitoneal injection, the expression of
Stra8 and of meiotic genes in germ cells was increased. The activation mechanism was suggested to be indirect via the activation of SMAD3 in pre-granulosa cells and the downregulation of CYP26B1 [
163].
2.3.2. Nutrient and Metabolic Factors
Nutrient restrictions and changes in metabolisms were recently proposed to act in concert with RA stimulation to activate the meiotic program [
167]. This is reminiscent of meiosis initiation in yeast, which mainly depends on nutrients and metabolism.
Autophagy is crucial for reserving energy in response to cellular stress conditions such as nutrient and oxygen starvation. In both fission and budding yeast, meiotic entry fails if autophagy is deficient [
170,
171]. Autophagy might contribute to degrade major meiotic entry inhibitors, thus allowing the cell to enter meiosis [
172]. However, in mammals, STRA8 represses autophagy by binding the promoter of NR1D1, which in turn leads to repressing its downstream target ULK1, an autophagy initiator, highlighting the requirement for the suppression of autophagy during meiosis initiation [
173].
This means that although the role of nutrient deprivation in meiosis is potentially conserved, it is yet to be fully understood in mammals. It can be hypothesized that the suppression of autophagy via STRA8 might be important in maintaining meiotic DSBs during prophase I, as autophagy plays a role in DNA damage repair. The master regulator of autophagy, mTORC1, was found to be crucial during meiotic onset. In fact, in both yeast and female Drosophila, the reduction in TORC1 expression in response to nutrient starvation was needed for the mitotic–meiotic switch [
174,
175]. In mammals, the suppression of mTORC1 activators is required for male mitotic arrest in PGCs, which could prime male germ cells for meiotic entry [
176].
This entry is adapted from the peer-reviewed paper 10.3390/ijms232012571