Your browser does not fully support modern features. Please upgrade for a smoother experience.
Please note this is an old version of this entry, which may differ significantly from the current revision.
Haploid plants with a doubled set of chromosomes (doubled haploid (DH)) significantly speed up the selection process by the fixation of genetic traits in each locus in the homozygous state within one generation. Doubled haploids are mainly attained by the formation of plants from the cultured gametophytic (haploid) tissues and cells in vitro, or by targeted reduction in the parent chromosome during intra- or interspecific hybridization. Since then, DH has become one of the most powerful tools to support various basic research studies, as well as applied research.
- doubled haploids
- in vivo
- BBM-like proteins
- Salmon system
1. Introduction
The process of crop breeding has been significantly accelerated through the ability to produce double haploid plants. Haploid plants are induced mostly by plant generation from cultivated gametophic tissues and cells using in vitro haploid techniques [1]. The challenge for breeding in the 21st century is to ensure the development of new varieties with the most valuable economic traits, in the shortest possible time, to meet the needs of agriculture in connection with increasing world population and global changes in climatic conditions [2]. The ability of plants to generate haploids with the further duplication of genome (doubled haploids) allows the significant speeding up of the agricultural cereals’ selection process. In recent decades, the doubled haploid system has appeared as an effective option to the traditional practice of pure line development [3][4][5]. This technology essentially samples segregating gametes from the germplasm of origin, often a biparental cross or a population, and produces fully homozygous lines in a single step. Both in vivo and in vitro systems can be applied to improve different crop lines, however, the in vivo technology has proved to be more reliable and efficient in large-scale production of doubled haploid lines [6][7].
The main advantage of the obtained doubled haploid pure lines for plant-breeders is the fixation of genetic traits in each locus in the homozygous state within one generation. This avoids the laborious traditional approach of numerous self-pollination repetitions or multiple backcrossings to obtain pure lines [8]. The genotype of the created doubled haploid line reproduces accurately and multiplies quickly, which helps plant-breeders to work with the number of traits they need using genetically homogeneous material, as well as to obtain outstanding selection efficiency at the early stage of the reproduction cycle. In numerous plant species, a viable haploid system is not available, or is only applicable for a limited genotype and is costly, although haploid cultures can be achieved by various methods [1][9].
2. Classic Doubled Haploids Production Technologies
The basis of cell technologies for obtaining doubled haploids of plant origin directly from gametophytic (haploid) cells present in various explants cultured in vitro are pistils, ovules, and isolated immature pollen (usually microspores; and less often, early binuclear pollen). Depending on the climate, regenerated crops inherit their haploid genome from male or female gametophytes, called maternal or paternal haploid, respectively.
Doubled haploid (DH) skill cuts plant breeding duration and offers homozygous pure lines with less effort [10]. It enhances proficiency in plant breeding programs by creating pure lines from DH crops, and enables selection of preferred genotypes. Mentioned valued lines have recently been applied in breeding programs and genetic studies [11]. Moreover, the DH technique helps to overcome inbreeding depression, and may be a possible substitute for creating homozygous lines [12].
Obtaining doubled haploids in vitro is performed by the methods of androgenesis (from male gametophytes) or gynogenesis (from female gametophytes). The androgenesis method consists of creating conditions for the transition of an anther culture, or a culture of isolated microspores, from the gametophytic development path to the sporophytic path through temperature shock and other cultivation features (pH, sucrose content) [13]. The method of gynogenesis involves obtaining doubled haploids from cultures of buds, ovaries, or unfertilized ovules.
Additionally, in vivo methods are used, including intra- or interspecific crosses. In vivo induction of maternal haploids can be induced through pollination with same species pollen, usually using classical inducers of haploidy. The basis of these methods is the elimination of one of the genomes, which usually occurs during early zygotic embryogenesis [14]. In intraspecific crosses, a haploinducer line is usually used. Pollination of maternal forms with pollen from haploinducer plants stimulates their parthenogenetic germ development. This method has been used successfully for many important crops. The result of using haploinducer lines depends not only on their ability for haploinduction, but also on the adaptability of the crossed forms to the geographic and climatic conditions of cultivation, the synchronization of flowering time, and other factors [15]. As an additional possibility of obtaining doubled haploids in vivo, the induction of parthenogenesis in plants after genetic manipulation or treatment with hormones has been reported. Haploidy can be obtained by pollination with pollen treated with ionizing radiation, certain chemicals, and high temperatures. In some techniques, when endosperm development does not occur, the “embryo rescue” method is needed. Subsequently, seedlings or haploid embryos spontaneously duplicate the genome or, more often, are processed by an agent that depolymerizes microtubules and leads to the disruption of their assembly in mitosis. This causes the doubling of the haploid genome and doubled haploid crop formation (Figure 1).
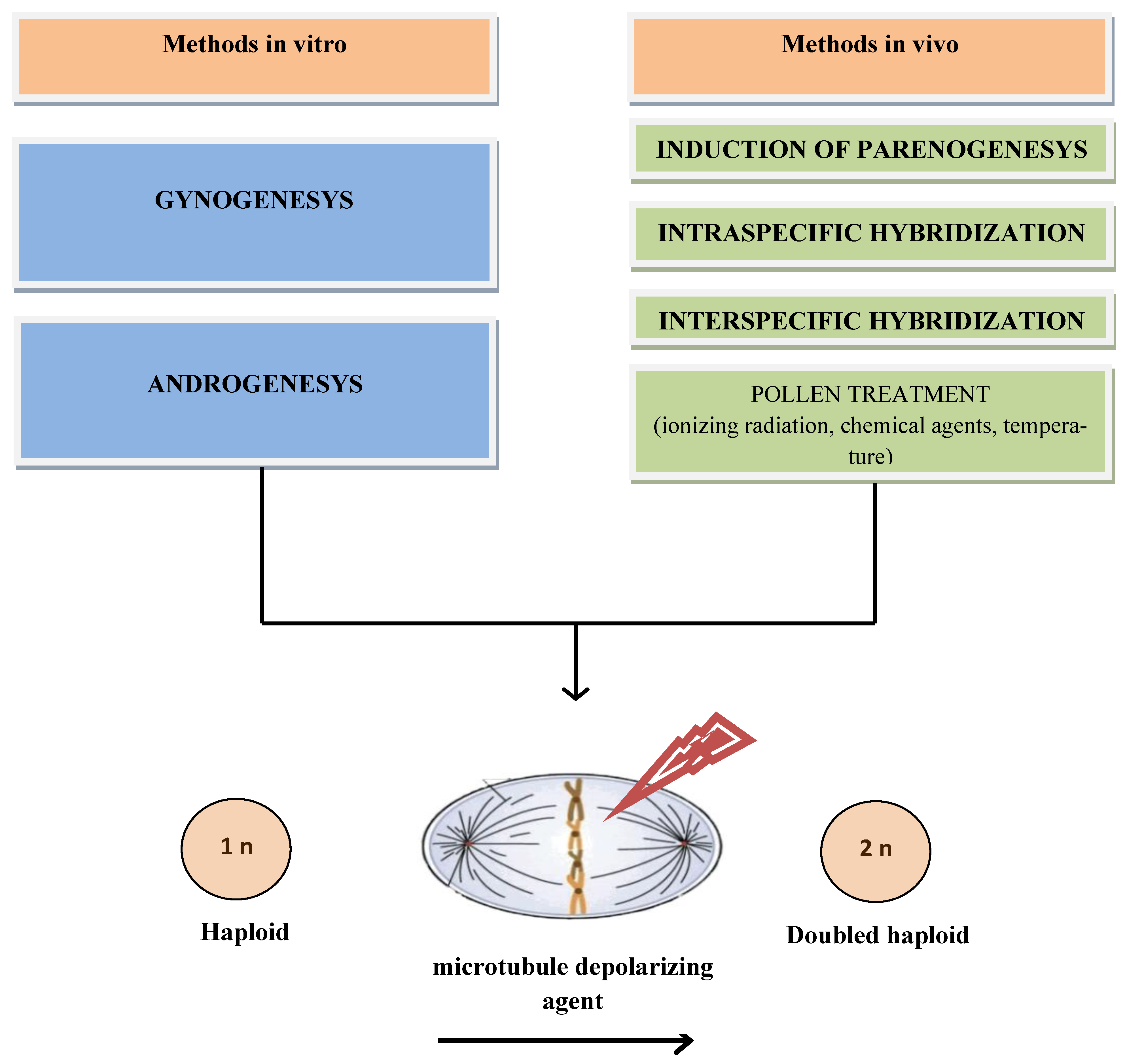
Figure 1. Representation of the methods of haploidy induction in plants.
3. The Recent Development for the Production of Doubled Haploids In Vivo and Their Fundamental Bases
Although obtaining doubled haploids can be achieved by various traditional methods, for many types of crops, obtaining viable doubled haploids is not yet available, or is usable only for limited genotype numbers and is economically disadvantageous [9][16][17][18]. Therefore, breeders are extremely interested in any developing techniques, as well as new haploidization principles.
The most recent studies on the mechanisms responsible for the formation of doubled haploids in vivo during intra- and interspecific hybridization are considered below, in particular, the perspective method of induction of parthenogenesis, and the method of intraspecific induction of doubled haploids in maize [5]. The latter method is used in practice and there are new data on its fundamental basis. The mechanisms that underlie the selective elimination of one of the genomes in early embryo cell division, and the methods of targeted manipulation of centromeres, are described in detail.
4. Production of Haploids by Induction of Parthenogenesis
4.1. BBM-Mediated Induction of Maternal Haploids
In recent years, the attention of researchers has been drawn to the genes associated with the processes of apomixis. Apomixis is asexual reproduction by plant seeds. It has been found in more than 300 plant species [19]. It is categorized into sporophytic and gametophytic types. The gametophytic type consists of apomeiosis and parthenogenesis processes. One of the main genes responsible for parthenogenesis is the PsASGR-BBML gene (PsASGR-BABY BOOM-like). It is similar to the BBM (BABY BOOM) genes, codes of transcription factors that were first discovered in Brassica napus. PsASGR-BBML has been found in Pennisetum squamulatum (pearl millet). This gene is located in the ASGR genomic region in multiple copies. The PsASGR-BBML gene was found to be expressed in the ovule prior to fertilization, in [20]. In this work, independent transgenic lines involving transgene gPsASGR-BBML were examined; the frequency of parthenogenesis ranged from 35 to 36%. In rice the most closely related gene to ASGR-BBML is OsBBM1, which is one of the rice BBM genes. The greatest expression of the gene was in the zygote rapidly activated de novo after fertilization, but there was no expression of this gene in the ovule [21]. It was reported that a wild-type rice OsBBM1 transgene under an Arabidopsis egg-cell-specific promoter (DD45) was able to initiate embryogenesis in cells of the rice egg without fertilization [22]. In maize, three homologs of BBM were recently shown to significantly increase expression after fertilization: ZmBBML1, ZmBBML2, and ZmLEC1 [23]. Conner et al. showed that the PsASGR-BBML transgene-induced parthenogenesis on rice and corn led to the formation of haploid embryos, with a frequency of 25 to 88%. The ability to create apomictic crops using genes identified from natural apomictic crops depends on the ability of these genes to function in crops. To obtain haploid embryos of the model plant Arabidopsis thaliana, both the endogenous PsASGR-BBML promoter from Pennisetum squamulatum, and the specific ovum promoter from Arabidopsis, were used. PsASGR-BBML was unable to induce measurable haploid seed growth in the Arabidopsis thaliana plant genetic model system [24]. However, the BBM (BABY BOOM) protein was able to induce somatic embryos in Brassica and Arabidopsis seedlings with ectopic overexpression [25]. The BBM protein has been submitted as a factor in the proliferation and morphogenesis of embryo cells. Additionally, ectopic expression of BBM1 in the egg cell was adequate for parthenogenesis. This gene is also expressed in sperm cells. Triple knockout of the genes BBM1, BBM2, and BBM3 caused embryo arrest and abortion, that were fully rescued with male-transmitted BBM1 [22]. Thereby, the induction of haploidy based on the use of BBM homologs is a perspective approach that requires thorough study.
4.2. The Salmon System in Wheat
The Salmon system is used in wheat production to examine the development of self-directed embryo growth at the cellular and molecular stage, as well as to induce complete apomixes within the parthenogenetic Salmon lines [26]. The first attempt to use parthenogenesis to induce haploids was in wheat. The “Salmon” lines were constructed in the 1990s. They were formed through the transfer of the nucleus of the sexual cultivar “Salmon” to the cytoplasm of the grass genus Aegilops. As a result, lines with capability for parthenogenesis were obtained [27]. The “Salmon system” of wheat was developed to apply in haploid production. Three isogenic alloplasmic lines with zygotic (aS) or autonomous, fertilization-independent (cS, kS) embryo development were identified. In these lines, the short arm of the 1B chromosome of wheat was replaced by the short arm of the 1R chromosome of rye, due to the exchange of two main genes of parthenogenesis control: Ptg (inducer) and Spg (suppressor). Further investigation revealed an α-tubulin polypeptide, named P115.1, expressed in the parthenogenetic lines from three days before anthesis [28]. However, it was unclear if this protein was the result of parthenogenesis or its cause. Kumlehn et al. demonstrated, in 2001, that parthenogenesis is an inherent feature of the isolated egg cell. Then, egg-cell-specific cDNA libraries were constructed with the following sequencing. In this research, some egg cell-specific candidate genes were obtained [29].
Among these candidates was the RWP-RK domain (RKD)-containing TFs. Their homologs were found in Arabidopsis thaliana, and were named AtRKDs [30]. Two of five AtRKDs were expressed in the egg cell and induced cell proliferation. Then it was revealed that RKD-TF genes are conserved among plants. Therefore, the analysis of the function of the RKD-TF homolog in Marchantia polymorpha (MpRKD) showed wide expression, developing egg cells and sperm precursor cells [31][32]. In MpRKD downregulated lines, cells at the base of the archegonium underwent cell divisions instead of transitioning to the resting egg stage. The MpRKD mutant had defects in egg and sperm differentiation [31][32]. Thus, RKD-TFs control the transition from the gametophytic to the sporophytic pathway of development, preventing the entry of the egg into mitosis in the absence of fertilization. Therefore, RKD-TFs are evolutionarily conserved regulators of germ cell differentiation, by suppressing parthenogenesis.
5. Creation of Haploids Using Intraspecific Hybridization in Maize
The technology of intraspecific crossing with the use of haploinducers was used in corn breeding for several decades. Haploid production with intraspecific hybridization is a key method in corn. It allows haploids to be obtained with a frequency of about 10%. For haploid induction in vivo, two approaches are mainly used: the first involves the ig1 gene (indeterminate gametophyte 1), and the second is based on derivatives of the Stock6 line.
5.1. IG1-System to Obtain Androgenic Haploids
Mutation in the ig1 gene, which results in approximately 3% of the frequency of haploid induction of paternal haploids, has been reported as a spontaneous mutation in the Wiscon-sin-23 (W23) line. This is a pure line that leads to approximately 3% of the frequency of haploid induction of paternal haploids. The ig1 gene is located on chromosome 3 and encodes the protein domain lateral organ boundaries (LOB) [33]. Proteins containing the LOB domain belong to a big family of transcription factors that are required for lateral organ development in higher crops [34]. In maize, pure line W23 ig1 mutant plants contain an insertion of the Hopscotch retrotransposon in the second exon of this gene, upstream of the site encoding the LOB domain [33]. Several phenotypes have been observed in viable ig1 mutants during the development of the female embryo sac. The embryo sacs were not divided into cells and contained an enhanced nuclei number due to violations of mitotic synchronization and abnormal behavior of microtubules; this can cause the formation of multiple embryos. Data showed that the function of wild-type IG1 is to facilitate the transition from proliferation to division in maize germ [33][35][36]. This hypothesis is supported with the fact that IG1 is the target of the ovum-secreted differentiation factor EAL1 in the signaling pathway [37]. However, the exact mechanisms of mutations in the ig1 gene which cause haploid induction, are yet to be investigated. It is known that the ig1 homolog in rice OsIG1 gene may be involved in the regulation of genes associated with the development of flower organs and the female gametophyte [38]. Additionally, it has been reported that the suppression of KNOX8 in embryo sacs by the ig1 gene indicates the existence of conservative gene regulation mechanisms [33].
The ability to produce paternal haploids has been applied in the breeding of maize to transfer chromosomes from a maize variety to the cytoplasm of another variety [39]. The technique is based in the carriage of the mutation in the ig1 gene in female plants. Thus, paternal haploids contain the female haploid cytoplasm and the haploid genome of the pollen donor. Maternal haploid offspring have also been attained using ig1 mutants, although the frequency of haploid induction was too low [40]. This system is used only in maize, and it is difficult to apply the IG1-system to other crops because of the unknown or exact mechanism of haploidization.
5.2. Maternal Haploid Induction Based on Stock6-System
The second approach used widely for the induction of haploidy in vivo in maize is based on the Stock6 line, with a high frequency of induction of haploidy. Stock6 is a pure line, with an induction of haploidy frequency of about 3% [41][42]. This line was used for the development of a genetic basis for new lines. Different types of hybrids were created through the crossing of the Stock6 line with the 1gl-mutant of the W23 line. They became the basis of many lines of haploinducers that are currently used (RWS, UH400, MHI, and PHI), which demonstrate a frequency of haploid induction of about 10% [43][44]. Numerous studies have been devoted to the search for genetic factors, in particular, quantitative trait loci (QTL), responsible for haploid induction on the mentioned lines.
In a 2008 study [45], the offspring obtained from the crossings of haploinducer line PK6 with a 6% haploidy induction, and non-inducer line DH99, were used for accurate genome mapping, which helped identify QTL associated with the induction of haploidy in chromosome 1, which was named ggi1. Recent studies showed that the ability to induce haploidy in lines derived from Stock6 is a complex quantitative trait controlled with different loci. Prigge et al. [44] identified a QTL that was obligatory for the induction of haploidy in chromosome 1 in the same region. It was named qhir1 [42][44]. During further verification, the locus qhirl was narrowed down from 50.3 Mbp to 243 kbp by accurate genome mapping. A new QTL important for haploidization induction was named qhir11 [46]. A big study using genome-wide sequencing was conducted to identify the subregion responsible for the induction of haploidy within the qhirl area. It used 53 haploinducer lines and 1482 non-haploinducer lines. This made it possible to identify a common QTL of 3.97 Mbp for all inductors, which was absent in all non-inductors and did not overlap with qhir11. The identified locus was named qhir12 [43].
The work of Nair et al. (2017) [2] was consistent with the study of Dong et al. (2013) [46] but not of Hu et al. (2016) [43]. Nair et al. (2017) illustrated the participation of the qhir1 subregions in the induction of haploidy [2]. Of the two qhir1 subregions, the region involving qhir11 had a desirable efficacy in the generation of maternal haploids regardless of the presence of the qhir12 allele, while the qhir12 allele of the haploinducer did not cause the induction of haploidy above the spontaneous frequency of the wild type. In addition, the region involving qhir11 caused segregation distortion and nuclear abortions, which are typical of haploinductor lines [2].
In 2017, several independent scientific papers by Gilles et al. [47][48], Kelliher et al. [49], and Liu et al. [50] research groups stated that the patatin-like phospholipase, which is a gene located at the qhir1 locus, was necessary for the induction of haploidy in lines derived from Stock6. Each of the research groups gave phospholipase a name: MTL, PLA1, and NLD, respectively. Hereinafter, this gene is referred as MTL.
MTL belongs to the superfamily of phospholipases A enzymes that catalyze the hydrolysis of phospholipid acyl groups to form free fatty acids and lysophospholipids. In the works of these authors, all studied haploinducer lines were characterized by a shortening of the protein product of the MTL gene. In addition, the insertion of four bp was found in the fourth exon of this gene. As a result of the insertion, the site was followed by twenty altered amino acids and a premature stop codon that shortened the protein by 29 amino acids. MTL mutants have been shown to induce haploids in vivo with typical side effects such as nuclear abortion and skewed chromosome segregation [47][48][49][50]. Using CRISPR/Cas9 gene editing, an average of 2% haploid induction and 9 to 14% nuclear abortions were indicated, and this was indicated to be the cause of the haploid induction phenotype [50]. Small deletions near the insertion site at 4 bp in lines derived from Stock6, induced with transcription-activator-like effector nuclease (TALEN), caused haploid induction in 4 to 13% (averaged 6.7%) of cases [49].
Gene expression researchers showed that MTL activity on corn is limited to mature pollen tubes and pollen grains [47]. Wild-type MTL, detected by the fluorophore contained at the C-terminus, is localized in the plasma membrane of spermium cells in germinated pollen tubes [47][48][49]. This finding suggests that the mutant variant is unstable in corn plant cells.
The mechanisms of the shortening of the MTL C-terminus which leads to the induction of haploidy are yet to be studied. Additionally, it is not completely clear how shortening affects MTL function. Gilles et al. suggested that the loss of membrane binding capacity in the mutant protein may play a vital role [47]. The authors studied the expression of the MTL gene in a model of Arabidopsis root epidermis with the constructed AtUBQ10 MTL-CITRINE promoter. They showed that the “wild” type protein was localized in the cytosol and plasma membrane, and its expression was also detected in small intracellular compartments. The shortened mutant version was not detected in the plasma membrane, being observed exclusively in the cytosol. Additional confirmation of this version was the fact that potential sites of S-palmitoylation or S-farnesylation at C423 were missing on the truncated protein [47].
One of the feasible consequences of reducing the C-terminus of phospholipase is the removal of a potential phosphorylation site, which could greatly affect the ability of protein to be activated during signaling in the cell. It was shown that the activity of the two closest homologues of the MTL gene in Arabidopsis, AtPLAIVA/PLP1 and AtPLAIVB/PLP5, depended on the phosphorylation of the C-terminus with calcium-dependent protein kinases (CDPKs) [51]. Thus, it is possible that the MTL gene product also needs phosphorylation at the C-terminus for correct signal transmission in cells during fertilization by a sperm-specific kinase, which is yet unknown.
Several suggestions were made to explain how changes in the MTL gene can lead to haploid induction and kernel abortion in haploinducer lines derived from Stock6. It may happen due to the inability to fertilize, while the ovule develops into a zygote (induction of haploidy through parthenogenesis). Haploinduction could also occur due to the development of endosperm from the central cell with the formation of a defective endosperm and subsequent kernel abortion, or by postzygotic elimination of the parental genome. However, the exact mechanism of haploinduction upon pollination by lines derived from Stock6, in particular by MTL mutants, remains unknown. In this regard, research is underway to study the processes of elimination of uniparental chromosomes [50][52][53][54] and single fertilization [55].
In addition to the qhir1 locus, which includes the MTL gene, another QTL, which is important for the induction of haploidy, was released in chromosome 9; it was named qhir8. This supported the idea that the qhir1 locus is modeled with other loci that are not capable of generating haploid induction on their own, and explained the observed 20% genotype dispersion [44][56][57].
The effect of the qhir8 locus on MTL function was studied by Liu et al. [56]. qhir8 was mapped in detail by analyzing the offspring obtained from the crossing of two haploinducers including a fixed qhir1 region: CAUHOI (haploid induction 2%), and UH400 (haploid induction 8%). The haploid induction of F2 plants, which are homozygous for qhir8 from the UH400 line, were found to be favorably greater than F2 plants, which are homozygous for qhir8 from the CAUHOI line. The average frequency of induction of haploidy in heterozygous F2 plants was between those in homozygous plants. These results confirmed that qhir8 can potentially increase the frequency of qhir1-mediated haploid induction [56].
The study of Zhong et al. allowed us to discover that the mutation of the ZmDMP gene in qhir8, increases and triggers haploid induction in maize [58]. There was a suggestion that in maize, the membrane protein ZmDMP is included in the male–female gamete interaction needed to attain correct double fertilization [59]. ZmDMP was identified by map-based cloning and further verified by CRISPR-Cas9-mediated knockout experiments. The ZmDMP gene was found to be highly expressed during the late stage of pollen development and localized to the plasma membrane; a single nucleotide change in ZmDMP led to a two- to three-fold enhancement in the haploidy induction. ZmDMP knockout triggered haploid induction with a 0.1 to 0.3% rate, and exhibited a greater ability to enhance haploidy induction by five–six-fold in the presence of MTL [58]. This synergetic effect implied that at least two distinct pathways exist in modern haploid inducer lines of maize [59].
Another recent research work focused on the characterization of positional candidate genes underlying qhir8. A strong candidate for qhir8, GRMZM2G435294 (MYO), was suppressed with RNAi. Analysis of crosses with these heterozygous RNAi transgenic lines for the rate of haploid induction showed that MYO silencing significantly increased the rate of haploid induction in the presence of qhir1, but only by 0.6% on average. However, reciprocal crosses revealed that the silencing of MYO causes male sterility [60]. Therefore, this gene is a candidate for further studies.
In 2018, an attempt was performed to induce haploidy in rice using the homologue of the MTL gene in this cereal—the OspPLAIIφ or OsMATL gene. This gene was subjected to site-directed modification using CRISPR/Cas9 technology. Mutants knocked out in this gene showed a reduced set of seeds and haploid induction with a frequency of up to 6% [61]. This allowed consideration of the possibility of transferring the approach using the MTL gene homologues, to other crops.
Further investigations revealed that loss-of-function mutations in ZmDMP-like genes AtDMP8 and AtDMP9 in Arabidopsis thaliana induced maternal haploids, with a haploid induction rate of 2.1 ± 1.1% on average. These findings indicated that mutations in DMP genes also trigger haploid induction in dicots. In the authors’ opinion, the conserved expression patterns and amino acid sequences of ZmDMP-like genes in dicots suggested that DMP mutations could be used to improve in vivo haploid induction systems in dicot plants [58].
Moreover, the genotype of the mother plant is also very important. Eder and Chalyk [62] examined the effect of source germplasm on the haploidy induction frequency of two lines by pollination of haploinducers MHI and M741H obtained from Stock6. Among the maternal genotypes tested were four European hard lines, eleven soft lines and five hard and soft hybrids. Maternal haploids were induced for all genotypes, albeit with different frequencies, and none of the genetic pools indicated an important advantage over the others [62]. For tropical corn, parental germplasm was indicated to be of great significance under different environmental conditions, resulting in differences in the frequency of induction of haploidy of 2.9–9.7% for the same pollinator [63]. Regarding accurate genome mapping, the maternal contribution to the induction of haploidy was found to be determined by the presence of two loci on chromosomes 1 and 3, called qmhir1 and qmhir2, respectively, which explained 14.7 and 8.4% of the genetic variation, respectively [64].
The research by De La Fuente et al. supported the use of germplasm with improved inducibility for breeding to develop an inducibility rate in germplasm that has low induction rates. The highest estimated inducibility rate was observed at 14.6% [65].
This entry is adapted from the peer-reviewed paper 10.3390/agriculture12101595
References
- Kalinowska, K.; Chamasb, S.; Unkel, K.; Demidov, D.; Lermontova, I.; Dresselhaus, T.; Kumlehn, J.; Dunemann, F. State-of-the-art and novel developments of in vivo haploid technologies. Theor. Appl. Genet. 2019, 132, 593–605.
- Nair, S.K.; Molenaar, W.; Melchinger, A.E.; Boddupalli, P.M.; Martinez, L.; Lopez, L.A.; Chaikam, V. Dissection of a major QTL qhir1conferring maternal haploid induction ability in maize. Theor. Appl. Genet. 2017, 130, 1113–1122.
- Yan, G.; Liu, H.; Wang, H.; Lu, Z.; Wang, Y.; Mullan, D.; Hamblin, J.; Liu, C. Accelerated generation of selfed pure line plants for gene identifcation and crop breeding. Front. Plant Sci. 2017, 8, 1786.
- Melchinger, A.E.; Böhm, J.; Utz, H.F.; Müller, J.; Munder, S.; Mauch, F.J. High-throughput precision phenotyping of the oil content of single seeds of various oilseed crops. Crop Sci. 2018, 58, 670–678.
- Meng, D.; Liu, C.; Chen, S.; Jin, W. Haploid induction and its application in maize breeding. Mol. Breed. 2021, 41, 20.
- Chaikam, V.; Molenaar, W.; Melchinger, A.; Boddupalli, P. Doubled haploid technology for line development in maize: Technical advances and prospects. Theor. Appl. Genet. 2019, 132, 3227–3243.
- Marimuthu, M.P.A.; Maruthachalam, R.; Bondada, R.; Kuppu, S.; Tan, E.H.; Britt, A. Epigenetically mismatched parental centromeres trigger genome elimination in hybrids. Sci. Adv. 2021, 7, 1–19.
- Brauner, P.C.; Schipprack, W.; Utz, H.F.; Bauer, E.; Mayer, M.; Schön, C.C.; Melchinger, A.E. Testcross performance of doubled haploid lines from European flint maize landraces are promising for broadening the genetic base of elite germplasm. Theor. Appl. Genet. 2019, 132, 1897–1908.
- Ishii, T.; Karimi-Ashtiyani, R.; Houben, A. Haploidization via chromosome elimination: Means and mechanisms. Annu. Rev. Plant Biol. 2016, 67, 421–438.
- Zhu, Y.C.; Sun, D.X.; Deng, Y.; An, G.L.; Li, W.H.; Si, W.J.; Liu, J.P.; Sun, X.P. Comparative transcriptome analysis of the effect of different heat shock periods on the unfertilized ovule in watermelon (Citrullus lanatus). J. Integr. Agric. 2020, 19, 528–540.
- Kurtar, E.S.; Balkaya, A. Production of in vitro haploid plants from in situ induced haploid embryos in winter squash (Cucurbita maxima Duchesne ex Lam.) via irradiated pollen. Plant Cell Tissue Org. Cult. 2010, 102, 267–277.
- Burlutsky, V.; Pronina, I.; Zavarykina, T.; Tulinova, E.; Tsygankova, N. Factors of the haploproducer method in the F1 hybrids system T. aestivum L.—Z. mays L. optimization. IOP Conf. Ser. Earth Environ. Sci. 2021, 723, 022087. (In Russain)
- Kanbar, O.Z.; Lantos, C.; Chege, P.K.; Kiss, E.; Pauk, J. Generation of doubled haploid lines from winter wheat (Triticum aestivum L.) breeding material using in vitro anther culture. Czech J. Genet. Plant Breed. 2020, 56, 150–158.
- Chen, C.; Xiao, Z.; Zhang, J.; Li, W.; Li, J.; Liu, C.; Chen, S. Development of In Vivo Haploid Inducer Lines for Screening Haploid Immature Embryos in Maize. Plants 2020, 9, 739.
- Andorf, C.; Beavis, W.D.; Hufford, M.; Smith, S.; Suza, W.P.; Wang, K.; Woodhouse, M.; Yu, J.; Lübberstedt, T. Technological advances in maize breeding: Past, present and future. Theor. Appl. Genet. 2019, 132, 817–849.
- Dwivedi, S.L.; Britt, A.B.; Tripathi, L.; Sharma, S.; Upadhyaya, H.D.; Ortiz, R. Haploids: Constraints and opportunities in plant breeding. Biotechnol. Adv. 2015, 33, 812–829.
- Kumlehn, J. Haploid technology. In Biotechnological Approaches to Barley Emprovement; Kumlehn, J., Stein, N., Eds.; Springer: Berlin/Heidelberg, Germany, 2014; Volume 69, pp. 379–392.
- Shen, Y.; Pan, G.; Lubberstedt, T. Haploid strategies for functionalvalidation of plant genes. Trends Biotechnol. 2015, 33, 611–620.
- Burlutsky, V.; Tulinova, A.; Tsygankova, V.; Pronina, I.; Zavarykina, T.; Zhiltsov, A.V.; Dedushev, I.; Romanova, E. Haploid albinism Hordeum vulgare L. In Modern Trends in Solving the Problems of the Agro-Industrial Complex Based on Innovative Technologies, Proceedings of the International Scientific and Practical Conference Dedicated to the 90th Anniversary of the Federal Research Center “Nemchinovka”, Volgograd, Russia, 23 April 2021; Voronova, S.I., Ed.; the Federal Research Center “Nemchinovka”: Volgograd, Russia, 2021; pp. 76–80. (In Russian)
- Conner, J.A.; Mookkan, M.; Huo, H.; Chae, K.; Ozias-Akins, P. A parthenogenesis gene of apomict origin elicits embryo formation from unfertilized eggs in a sexual plant. Proc. Natl. Acad. Sci. USA 2015, 112, 11205–11210.
- Anderson, S.N.; Johnson, C.S.; Chesnut, J.; Jones, D.S.; Khanday, I.; Woodhouse, M.; Li, C.; Conrad, L.J.; Russell, S.D.; Sundaresan, V. The zygotic transition is initiated in unicellular plant zygotes with asymmetric activation of parental genomes. Dev. Cell 2017, 43, 349–358.
- Khanday, I.; Skinner, D.; Yang, B.; Mercier, R.; Sundaresan, V. A male-expressed rice embryogenic trigger redirected for asexual propagation through seeds. Nature 2019, 565, 91–95.
- Chen, J.; Strieder, N.; Krohn, N.G.; Cyprys, P.; Sprunck, S.; Engelmann, J.C.; Dresselhaus, T. Zygotic genome activation occurs shortly after fertilization in maize. Plant Cell 2017, 29, 2106–2125.
- Conner, J.A.; Podio, M.; Ozias-Akins, P. Haploid embryo production in rice and maize induced by PsASGR-BBML transgenes. Plant Reprod. 2017, 30, 41–52.
- Boutilier, K.; Offringa, R.; Sharma, V.K.; Kieft, H.; Ouellet, T.; Zhang, L.; Hattori, J.; Liu, C.M.; van Lammeren, A.A.; Miki, B.L.; et al. Ectopic expression of BABY BOOM triggers a conversion from vegetative to embryonic growth. Plant Cell 2002, 14, 1737–1749.
- Ghadamkheir, M.; Vladimirovich, K.P.; Orujov, E.; Bayat, M.; Madumarov, M.M.; Avdotyin, V.; Zargar, M. Influence of sulfur fertilization on infection of wheat Take-all disease caused by the fungus Gaeumannomyces graminis var. tritici. Res. Crops 2020, 21, 627–633.
- Tsunewaki, K.; Mukai, Y. Wheat haploids through the Salmon method. In Biotechnology Agriculture Forestry; Bajaj, Y.P.S., Ed.; Springer: Berlin/Heidelberg, Germany, 1990; Volume 13, pp. 460–478.
- Matzk, F.; Meyer, H.M.; Bäumlein, H.; Balzer, H.J.; Schubert, I. A novel approach to the analysis of the initiation of embryo development in Gramineae. Sex. Plant Reprod. 1995, 8, 266–272.
- Kumlehn, J.; Kirik, V.; Czihal, A.; Altschmied, L.; Matzk, F.; Lörz, H.; Bäumlein, H. Parthenogenetic egg cells of wheat: Cellular and molecular studies. Sex. Plant Reprod. 2001, 14, 239–243.
- Kőszegi, D.; Johnston, A.J.; Rutten, T.; Czihal, A.; Altschmied, L.; Kumlehn, J.; Wüst, S.E.J.; Kirioukhova, O.; Gheyselinck, J.; Grossniklaus, U.; et al. Members of the RKD transcription factor family induce an egg cell-like gene expression program. Plant J. 2011, 67, 280–291.
- Koi, S.; Hisanaga, T.; Sato, K.; Shimamura, M.; Yamato, K.T.; Ishizaki, K.; Kohchi, T.; Nakajima, K. An evolutionarily conserved plant RKD factor controls germ cell differentiation. Curr. Biol. 2016, 26, 1–7.
- Rövekamp, M.; Bowman, J.L.; Grossniklaus, U. Marchantia MpRKD regulates the gametophyte-sporophyte transition by keeping egg cells quiescent in the absence of fertilization. Curr. Biol. 2016, 26, 1–8.
- Evans, M.M. The indeterminate gametophyte1 gene of maize encodes a LOB domain protein required for embryo sac and leaf development. Plant Cell 2007, 19, 46–62.
- Husbands, A.; Bell, E.M.; Shuai, B.; Smith, H.M.; Springer, P.S. Lateral Organ Boundaries defines a new family of DNA binding transcription factors and can interact with specific bHLH proteins. Nucleic Acids Res. 2007, 35, 6663–6671.
- Guo, F.; Huang, B.Q.; Han, Y.; Zee, S.Y. Fertilization in maize indeterminategametophyte1 mutant. Protoplasma 2004, 223, 111–120.
- Huang, B.Q.; Sheridan, W.F. Embryo sac development in the maize indeterminate gametophyte1 mutant: Abnormal nuclear behavior and defective microtubule organization. Plant Cell 1996, 8, 1391–1407.
- Krohn, N.G.; Lausser, A.; Juranic, M.; Dresselhaus, T. Egg cell signalingby the secreted peptide ZmEAL1 controls antipodal cellfate. Dev. Cell 2012, 23, 219–225.
- Zhang, J.; Tang, W.; Huang, Y.; Niu, X.; Zhao, Y.; Han, Y.; Liu, Y. Down-regulation of an LBD-like gene, OsIG1, leads to the occurrence of unusual double ovules and developmental abnormalities of various floral organs and megagametophyte in rice. J. Exp. Bot. 2015, 66, 99–112.
- Ren, J.; Wu, P.; Trampe, B.; Tian, X.; Lubberstedt, T.; Chen, S. Noveltechnologies in doubled haploid line development. Plant Biotechnol. J. 2017, 15, 1361–1370.
- Kermicle, J.L. Androgenesis conditioned by a mutation in maize. Science 1969, 166, 1422–1424.
- Coe, E.H. A line of maize with high haploid frequency. Am. Nat. 1959, 93, 381–382.
- Li, Y.; Lin, Z.; Yue, Y.; Zhao, H.; Fei, X.; Liu, C.; Chen, S.; Lai, J.; Song, W. Loss-of-function alleles of ZmPLD3 cause haploid induction in maize. Nat. Plants 2021, 7, 1579–1588.
- Hu, H.; Schrag, T.A.; Peis, R.; Unterseer, S.; Schipprack, W.; Chen, S.; Lai, J.; Yan, J.; Prasanna, B.M.; Nair, S.K.; et al. The genetic basis of haploid induction in maize identified with a novel genome-wide association method. Genetics 2016, 202, 1267–1276.
- Prigge, V.; Xu, X.; Li, L.; Babu, R.; Chen, S.; Atlin, G.N.; Melchinger, A.E. New insights into the genetics of in vivo induction of maternal haploids, the backbone of doubled haploid technologyin maize. Genetics 2012, 190, 781–793.
- Barret, P.; Brinkmann, M.; Beckert, M. A major locus expressed in the male gametophyte with incomplete penetrance is responsible for in situ gynogenesis in maize. Theor. Appl. Genet. 2008, 117, 581–594.
- Dong, X.; Xu, X.; Miao, J.; Li, L.; Zhang, D.; Mi, X.; Liu, C.; Tian, X.; Melchinger, A.E.; Chen, S. Fine mapping of qhir1 influencing in vivo haploid induction in maize. Theor. Appl. Genet. 2013, 126, 1713–1720.
- Gilles, L.M.; Khaled, A.; Laffaire, J.B.; Chaignon, S.; Gendrot, G.; Laplaige, J.; Bergès, H.; Beydon, G.; Bayle, V.; Barret, P.; et al. Loss of pollen-specific phospholipase NOT LIKE DAD triggers gynogenesis in maize. EMBO J. 2017, 36, 707–717.
- Gilles, L.M.; Martin, J.P.; Rogowsky, P.M.; Widiez, T. Haploidinduction in plants. Curr. Biol. 2017, 27, 1095–1097.
- Kelliher, T.; Starr, D.; Richbourg, L.; Chintamanani, S.; Delzer, B.; Nuccio, M.L.; Green, J.; Chen, Z.; McCuiston, J.; Wang, W.; et al. MATRILINEAL, a sperm-specific phospholipase, triggers maize haploid induction. Nature 2017, 542, 105–109.
- Liu, C.; Li, X.; Meng, D.; Zhong, Y.; Chen, C.; Dong, X.; Xu, X.; Chen, B.; Li, W.; Li, L.; et al. A 4-bp insertion at ZmPLA1 encoding aputative phospholipase A generates haploid induction in maize. Mol. Plant 2017, 10, 520–522.
- Rietz, S.; Dermendjiev, G.; Oppermann, E.; Tafesse, F.G.; Effendi, Y.; Holk, A.; Parker, J.E.; Teige, M.; Scherer, G.F. Roles of Arabidopsis patatin-related phospholipases a in root development arerelated to auxin responses and phosphate deficiency. Mol. Plant 2010, 3, 524–538.
- Li, L.; Xu, X.; Jin, W.; Chen, S. Morphological and molecular evidences for DNA introgression in haploid induction via a high oilinducer CAUHOI in maize. Planta 2009, 230, 367–376.
- Zhang, Z.; Qiu, F.; Liu, Y.; Ma, K.; Li, Z.; Xu, S. Chromosome elimination and in vivo haploid production induced by Stock6-derived inducer line in maize (Zea mays L.). Plant Cell Rep. 2008, 27, 1851–1860.
- Zhao, X.; Xu, X.; Xie, H.; Chen, S.; Jin, W. Fertilization and uniparental chromosome elimination during crosses with maize haploid inducers. Plant Physiol. 2013, 163, 721–731.
- Tian, X.; Qin, Y.; Chen, B.; Liu, C.; Wang, L.; Li, X.; Dong, X.; Liu, L.; Chen, S. Hetero-fertilization along with failed egg-sperm cell fusion supports single fertilization involved in in vivo haploidinduction in maize. J. Exp. Bot. 2018, 69, 4689–4701.
- Liu, C.; Li, W.; Zhong, Y.; Dong, X.; Hu, H.; Tian, X.; Wang, L.; Chen, B.; Chen, C.; Melchinger, A.E.; et al. Fine mapping of qhir8 affecting in vivo haploid induction in maize. Theor. Appl. Genet. 2015, 128, 2507–2515.
- Li, J.; Cheng, D.; Guo, S.; Yang, Z.; Chen, M.; Chen, C.; Jiao, Y.; Li, W.; Liu, C.; Zhong, Y.; et al. Genomic selection to optimize doubled haploid-based hybrid breeding in maize. bioRxiv 2010.
- Zhong, Y.; Liu, C.; Qi, X.; Jiao, Y.; Wang, D.; Wang, Y.; Liu, Z.; Chen, C.; Chen, B.; Tian, X.; et al. Mutation of ZmDMP enhances haploid induction in maize. Nat. Plants 2019, 5, 575–580.
- Jacquier, N.M.A.; Gilles, L.M.; Pyott, D.E.; Martinant, J.-P.; Rogowsky, P.M.; Widiez, T. Puzzling out plant reproduction by haploid induction for innovations in plant breeding. Nat. Plants 2020, 6, 610–619.
- Vanous, K.; Lübberstedt, T.; Ibrahim, R.; Frei, U.K. MYO, a Candidate Gene for Haploid Induction in Maize Causes Male Sterility. Plants 2020, 9, 773.
- Yao, L.; Zhang, Y.; Liu, C.; Liu, Y.; Wang, Y.; Liang, D.; Liu, J.; Sahoo, G.; Kelliher, T. OsMATL mutation induces haploid seed formation in indica rice. Nat. Plants 2018, 4, 530–533.
- Eder, J.; Chalyk, S. In vivo haploid induction in maize. Theor. Appl. Genet. 2002, 104, 703–708.
- Kebede, A.Z.; Dhillon, B.S.; Schipprack, W.; Araus, J.L.; Bänziger, M.; Semagn, K.; Alvarado, G.; Melchinger, A.E. Effect of source germplasm and season on the in vivo haploid induction rate intropical maize. Euphytica 2011, 180, 219–226.
- Wu, P.; Li, H.; Ren, J.; Chen, S. Mapping of maternal QTLs for in vivo haploid induction rate in maize (Zea mays L.). Euphytica 2014, 196, 413–421.
- De La Fuente, G.N.; Frei, U.K.; Trampe, B.; Nettleton, D.; Zhang, W.; Lübberstedt, T. A diallel analysis of a maize donor population response to in vivo maternal haploid induction: I. inducibility. Crop Sci. 2018, 58, 1830–1837.
This entry is offline, you can click here to edit this entry!