Eurypterus (/jʊəˈrɪptərəs/ yoo-RIP-tər-əs) is an extinct genus of eurypterid, a group of organisms commonly called "sea scorpions". The genus lived during the Silurian period, from around 432 to 418 million years ago. Eurypterus is by far the most well-studied and well-known eurypterid. Eurypterus fossil specimens probably represent more than 95% of all known eurypterid specimens. There are fifteen species belonging to the genus Eurypterus, the most common of which is E. remipes, the first eurypterid fossil discovered and the state fossil of New York. Members of Eurypterus averaged at about 13 to 23 cm (5 to 9 in) in length, but the largest individual discovered was estimated to be 60 cm (24 in) long. They all possessed spine-bearing appendages and a large paddle they used for swimming. They were generalist species, equally likely to engage in predation or scavenging.
- silurian
- eurypterid
- eurypterus
1. Discovery

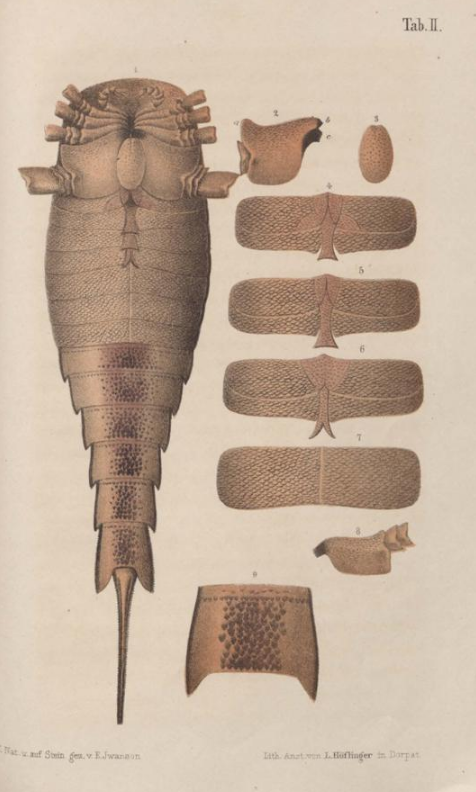
The first fossil of Eurypterus was found in 1818 by S. L. Mitchill, a fossil collector. It was recovered from the Bertie Formation of New York (near Westmoreland, Oneida County). Mitchill interpreted the appendages on the carapace as barbels arising from the mouth.[1] He consequently identified the fossil as a catfish of the genus Silurus.[2][3]
It was only after seven years, in 1825, that the American zoologist James Ellsworth De Kay identified the fossil correctly as an arthropod.[4] He named it Eurypterus remipes and established the genus Eurypterus in the process. The name means 'wide wing' or 'broad paddle', referring to the swimming legs, from Greek εὐρύς (eurús 'wide') and πτερόν (pteron 'wing').[1]
However, De Kay thought Eurypterus was a branchiopod (a group of crustaceans which include fairy shrimps and water fleas).[4] Soon after, Eurypterus lacustris was also discovered in New York in 1835 by the paleontologist Richard Harlan. Another species was discovered in Estonia in 1858 by Jan Nieszkowski. He considered it to be of the same species as the first discovery (E. remipes); it is now known as E. tetragonophthalmus.[1] These specimens from Estonia are often of extraordinary quality, retaining the actual cuticle of their exoskeletons. In 1898, the Swedish paleontologist Gerhard Holm separated these fossils from the bedrock with acids. Holm was then able to examine the almost perfectly preserved fragments under a microscope. His remarkable study led to the modern breakthrough on eurypterid morphology.[2]
More fossils were recovered in great abundance in New York in the 19th century, and elsewhere in eastern Eurasia and North America. Today, Eurypterus remains one of the most commonly found and best known eurypterid genera, comprising more than 95% of all known eurypterid fossils.[5]
E. remipes was designated the New York State Fossil by the then Governor Mario Cuomo in 1984.[6]
2. Description
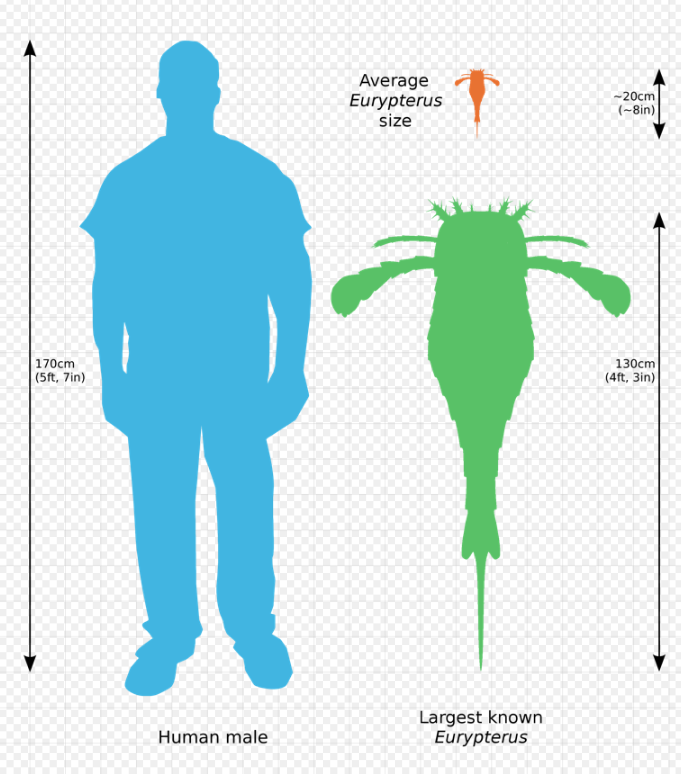
The largest arthropods to have ever existed were eurypterids. The largest known species (Jaekelopterus rhenaniae) reached up to 2.5 m (8.2 ft) in length, about the size of a crocodile.[7] Species of Eurypterus, however, were much smaller.
E. remipes are usually between 13 to 20 cm (5 to 8 in) in length. E. lacustris average at larger sizes at 15 to 23 cm (6 to 9 in) in length.[8] However, a single telson (the posteriormost division of the body) of a specimen of this species reaches this length, being almost 15 cm (5.9 in) long and indicating a specimen of more than half a meter of length.[9] The largest specimen of E. remipes ever found was 1.3 m (4.3 ft) long, currently on display at the Paleontological Research Institution of New York.[10]
Eurypterus fossils often occur in similar sizes in a given area. This may be a result of the fossils being "sorted" into windrows as they were being deposited in shallow waters by storms and wave action.
The Eurypterus body is broadly divided into two parts: the prosoma and the opisthosoma (in turn divided into the mesosoma and the metasoma).[1][11]
The prosoma is the forward part of the body, it is actually composed of six segments fused together to form the head and the thorax.[11] It contains the semicircular to subrectangular platelike carapace. On the dorsal side of the latter are two large crescent-shaped compound eyes.[12] They also possessed two smaller light-sensitive simple eyes (the median ocelli) near the center of the carapace on a small elevation (known as the ocellar mound).[13] Underneath the carapace is the mouth and six appendages, usually referred to in Roman numerals I-VI. Each appendage in turn is composed of nine segments (known as podomeres) labeled in Arabic numerals 1–9. The first segments which connect the appendages to the body are known as the coxa (plural coxae).[13]
The first pair (Appendage I) are the chelicerae, small pincer-like arms used for tearing food apart (mastication) during feeding. After the chelicerae are three pairs of short legs (Appendages II, III, and IV). They are spiniferous, with predominantly two spines on each podomere and with the tipmost segment having a single spine. The last two segments are often indistinguishable and give the appearance of a single segment having three spines.[14] They are used both for walking and for food capture. The next pair (Appendage V) is the most leg-like of all appendages, longer than the first three pairs and are mostly spineless except at the tipmost segments. The last pair (Appendage VI) are two broad paddle-like legs used for swimming.[1] The coxae of Appendage VI are broad and flat, resembling an 'ear'.[13]
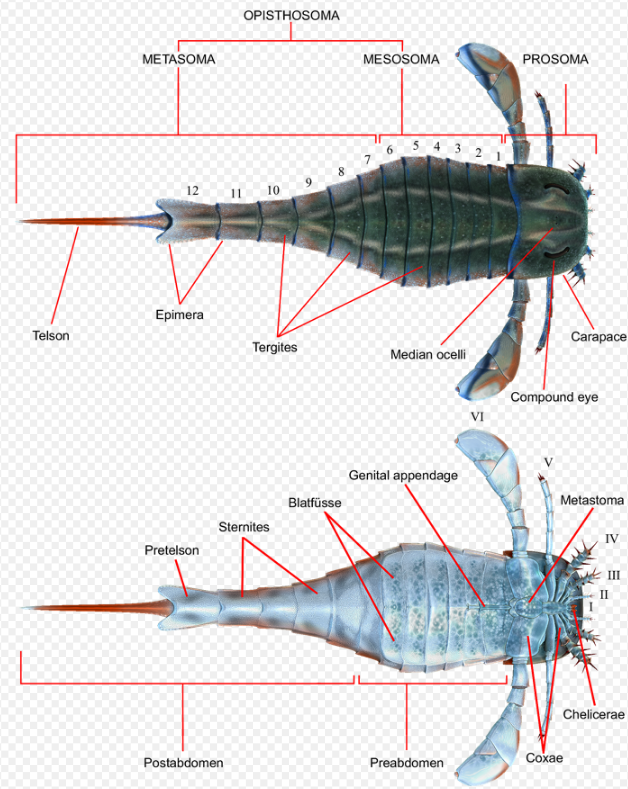
The ophisthosoma (the abdomen) is composed of 12 segments, each consisting of a fused upper plate (tergite) and bottom plate (sternite).[15] It is further subdivided in two ways.
Based on the width and structure of each segment, they can be divided into the broad preabdomen (segments 1 to 7) and the narrow postabdomen (segments 8 to 12).[16] The preabdomen is the broader segments of the anterior portion of the ophisthosoma while the postabdomen are the last five segments of the Eurypterus body. Each of the segments of the postabdomen contain lateral flattened protrusions known as the epimera with the exception of the last needle-like (styliform) part of the body known as the telson. The segment immediately preceding the telson (which also has the largest epimera of the postabdomen) is known as the pretelson.[13]
An alternative way to divide the ophisthosoma is by function. It can also be divided into the mesosoma (segments 1 to 6), and the metasoma (segments 7 to 12).[12][13][17] The mesosoma contains the gills and reproductive organs of Eurypterus. Its ventral segments are overlaid by appendage-derived plates known as Blatfüsse (singular Blatfuss, German for "sheet foot"). Protected within which are the branchial chambers which contain the respiratory organs of Eurypterus.[18][19] The metasoma, meanwhile, do not possess Blatfüsse.[17]
Some authors incorrectly use mesosoma and preabdomen interchangeably, as with metasoma and postabdomen.
The main respiratory organs of Eurypterus were what seems to be book gills, located in branchial chambers within the segments of the mesosoma. They may have been used for underwater respiration.[11] They are composed of several layers of thin tissue stacked in such a way as to resemble the pages of a book, hence the name. In addition, they also possessed five pairs of oval-shaped areas covered with microscopic projections on the ceiling of the second branchial chambers within the mesosoma, immediately below the gill tracts. These areas are known as Kiemenplatten (or gill-tracts, though the former term is preferred). They are unique to eurypterids.[19][20]
Eurypterus are sexually dimorphic. On the bottom side of the first two segments of the mesosoma are central appendages used for reproduction. In females, they are long and narrow. In the males they are very short.[21] A minority of authors, however, assume the reverse: longer genital appendage for males, shorter for females.[22]
The exoskeleton of Eurypterus is often covered with small outgrowths known as ornamentation. They include pustules (small protrusions), scales, and striations.[15] They vary by species and are used for identification. For more detailed diagnostic descriptions of each species under Eurypterus, see sections below.[23]
3. Classification
The genus Eurypterus belongs to the family Eurypteridae. They are classified under the superfamily Eurypteroidea, suborder Eurypterina, order Eurypterida, and the subphylum Chelicerata.[24] Until recently, eurypterids were thought to belong to the class Merostomata along with order Xiphosura. It is now believed that eurypterids are a sister group to Arachnida, closer to scorpions and spiders than to horseshoe crabs.[18][25][26]
Eurypterus was the first recognized taxon of eurypterids and is the most common. As a consequence, nearly every remotely similar eurypterid in the 19th century was classified under the genus (except for the distinctive members of the family Pterygotidae and Stylonuridae). The genus was eventually split into several genera as the science of taxonomy developed.[4]
In 1958, several species distinguishable by closer placed eyes and spines on their swimming legs were split off into the separate genus Erieopterus by Erik Kjellesvig-Waering.[4] Another split was proposed by Leif Størmer in 1973 when he reclassified some Eurypterus to Baltoeurypterus based on the size of some of the last segments of their swimming legs. O. Erik Tetlie in 2006 deemed these differences too insignificant to justify a separate genus. He merged Baltoeurypterus back into Eurypterus. It is now believed that the minor variations described by Størmer are simply the differences found in adults and juveniles within a species.[14]

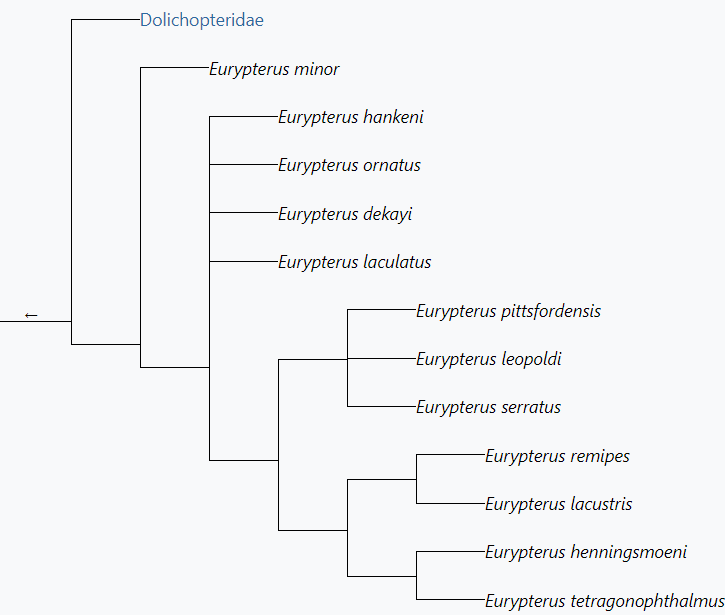
The genus Eurypterus derives from E. minor, the oldest known species from the Llandovery of Scotland. E. minor is believed to have diverged from Dolichopterus macrocheirus sometime in the Llandovery. The following is the phylogenetic tree of Eurypterus based on phylogenetic studies by O. Erik Tetlie in 2006. Some species are not represented.[23]
4. Species




Species belonging to the genus, their diagnostic descriptions, synonyms (if present), and distribution are as follows:[24]
Eurypterus De Kay, 1825
- ?Eurypterus cephalaspis Salter, 1856 – Silurian, England
- Eurypterus dekayi Hall, 1859 – Silurian, United States & Canada
-
- No raised scales on the posterior margin of the carapace or of the three front-most tergites. The rest of the tergites each have four raised scales. Four to six spines on each podomere of Appendages III and IV. Pretelson has large, rounded epimera without ornamentation on the margins. The species is very similar to E. laculatus. The species is named after James Ellsworth De Kay. Specimens recovered from New York and Ontario.[23]
- Eurypterus flintstonensis Swartz, 1923 – Silurian, USA
-
- Probably a synonym of E. remipes or E. lacustris. Probably named after Flintstone, Georgia (?). Specimen recovered from eastern United States.[23]
- Eurypterus hankeni Tetlie, 2006 – Silurian, Norway
-
- Small Eurypterus species, averaging at 10 to 15 centimetres (3.9 to 5.9 in) long. The largest specimen found is about 20 to 25 centimetres (7.9 to 9.8 in) in length. They can be distinguished by pustules and six scales at the rear margin of their carapaces. Appendages I to IV has two spines on each podomere. The postabdomen have small epimera. The pretelson has long pointed epimera. Telson has striations near its attachment to the pretelson. The species is named after Norwegian paleontologist Nils-Martin Hanken, of the University of Tromsø. Found in the Steinsfjorden Formation of Ringerike, Norway.[23][24]
- Eurypterus henningsmoeni Tetlie, 2002 – Silurian, Norway
-
- Eurypterus with broad paddles and metastoma. Postabdomen has small epimera. Pretelson has large rounded epimera with imbricate scales (overlapping, similar to fish scales). It is very similar and closely related to E. tetragonophthalmus. The species was named after the Norwegian paleontologist Gunnar Henningsmoen. Found in Bærum, Norway.[23][24]
- Eurypterus laculatus Kjellesvig-Waering, 1958 – Silurian, USA & Canada
-
- The visual area of the compound eyes of this species are surrounded by depressions. The ocelli and the ocellar mound are small. No pustules or raised scales on the carapace or the first tergite. It is probably closely related to E. dekayi. Its specific epithet means 'four-cornered', from Latin laculatus ('four-cornered, checkered'). Found in New York and Ontario.[23]
- Eurypterus lacustris Harlan, 1834 – Silurian, USA & Canada
-
-
- = Eurypterus pachycheirus Hall, 1859 – Silurian, USA & Canada
- = Eurypterus robustus Hall, 1859 – Silurian, USA & Canada
- One of the two most common Eurypterus fossils found. It is very similar to E. remipes and often found in the same localities, but the eyes are placed at a more posterior position on the carapace of E. lacustris. It is also slightly larger with a slightly narrower metastoma. Its status as a distinct species was once disputed before diagnostic analysis by Tollerton in 1993. Its specific name means 'from a lake', from Latin lacus ('lake'). Found in New York and Ontario.[23]
-
- Eurypterus leopoldi Tetlie, 2006 – Silurian, Canada
-
- Frontmost tergite is reduced. Metasoma is rhombiovate in shape with tooth-like projections at the anterior part. The pretelson has serrated edges. the epimera are large, semi-angular with angular striations. The telson is styliform with large angular striations interspersed among smaller more numerous striations. The species is named after Port Leopold and the Leopold Formation where they were collected. Found in the Leopold Formation of Somerset Island, Canada.[23]
- Eurypterus megalops Clarke & Ruedemann, 1912 – Silurian, USA
-
- Specific name means "large eye", from Greek μέγας (megas 'big or large') and ὤψ (ōps 'eye'). Discovered in New York, United States.[24]
- ?Eurypterus minor Laurie, 1899 – Silurian, Scotland
-
- Small Eurypterus with large pustules on the carapace and abdomen. Does not possess the scale ornamentation found in other species of Eurypterus. It is the earliest known species of Eurypterus. They have large palpebral lobes (part of "cheeks" of the carapace adjacent to the compound eyes), making it easy to mistake their eyes for being oval. This enlargement is more typical of the genus Dolichopterus and it may actually belong to Dolichopteridae.[5] The specific name means 'smaller', from Latin minor. Found in the Reservoir Formation of Pentland Hills, Scotland.[23]
- Eurypterus ornatus Leutze, 1958 – Silurian, USA
-
- Ornamentation of pustules on the entire surface of the carapace and at least the first tergite. Does not possess raised scales. Its specific name means 'adorned', from Latin ōrnātus ('adorned, ornate'). Recovered from Fayette, Ohio.[23]
- Eurypterus pittsfordensis Sarle, 1903 – Silurian, USA
-
- The posterior margin of the carapace has three raised scales. Appendages II to IV has two spines per podomere. The metastoma is rhomboid in shape with a deep notch at the front part. The postabdomen has serrated fringes at the middle with small angular epimera at the sides. The pretelson has large, semiangular epimera with angular striations at the margins. The telson is styliform with sparse angular striations at the margins. The name of the species comes from its place of discovery – the Salina shale formations of Pittsford, New York.[23]
- Eurypterus quebecensis Kjellesvig-Waering, 1958 – Silurian, Canada
-
- Has six raised scales on the posterior margin of the carapace but does not possess pustule ornamentation. It is named after the location it was recovered from Quebec, Canada.
- Eurypterus remipes DeKay, 1825 – Silurian, USA, Canada
-
-
- = Carcinosoma trigona (Ruedemann, 1916) – Silurian, USA
- The most common Eurypterus species. Has four raised scales at the posterior margin of the carapace. Appendages I to IV has two spines on each podomere. Postabdomen has small epimera. Pretelson has small, semiangular epimera with imbricate scale ornamentation at the margins. The telson has serrated margins along most of its length. It is very similar to E. lacustris and can often only be distinguished by the position of the eyes. The specific name means 'oar-foot', from Latin rēmus ('oar') and pes ('foot'). Found in New York and Ontario.[23]
-
- Eurypterus serratus (Jones & Woodward, 1888) – Silurian, Sweden
-
- Similar to E. pittsfordensis and E. leopoldi but can be distinguished by the dense angular striations on their styliform telson. The specific name means 'serrated', from Latin serrātus ('sawn [into pieces]'). Originally discovered from Gotland, Sweden.[23]
- Eurypterus tetragonophthalmus Fischer, 1839 – Silurian, Ukraine & Estonia
-
-
- = Eurypterus fischeri Eichwald, 1854 – Silurian, Ukraine
- = Eurypterus fischeri var. rectangularis Schmidt, 1883 – Silurian, Estonia
- Four raised scales on the posterior margin of the carapce. Appendages II to IV each have two spines on each podomere. Postabdomen has small epimera. The pretelson has large, rounded epimera with imbricate scale ornamentation at the margins. Telson has imbricate scale ornamentations at the margins of the base which become serrations towards the tip. The specific name means 'four-edged eye', from Greek τέσσαρες (tessares 'four'), γωνία (gōnia 'angle'), and ὀφθαλμός (ophthalmos 'eye'). Found in the Rootsiküla Formation of Saaremaa (Ösel), Estonia, with additional discoveries in Ukraine, Norway, and possibly Moldova and Romania.[23]
-
The list does not include the large number of fossils previously classified under Eurypterus. Most of them are now reclassified to other genera, identified as other animals (like crustaceans) or pseudofossils, or remains of doubtful placement. Classification is based on Dunlop et al.(2011).[24]
5. Paleobiology
Eurypterus belongs to the suborder Eurypterina, eurypterids in which the sixth appendage had developed a broad swimming paddle remarkably similar to that of the modern-day swimming crab. Modeling studies on Eurypterus swimming behavior suggest that they utilized a drag-based rowing type of locomotion where appendages moved synchronously in near-horizontal planes.[22] The paddle blades are almost vertically oriented on the backward and down stroke, pushing the animal forward and lifting it up. The blades are then oriented horizontally on the recovery stroke to slash through the water without pushing the animal back. This type of swimming is exhibited by crabs and water beetles.[27]
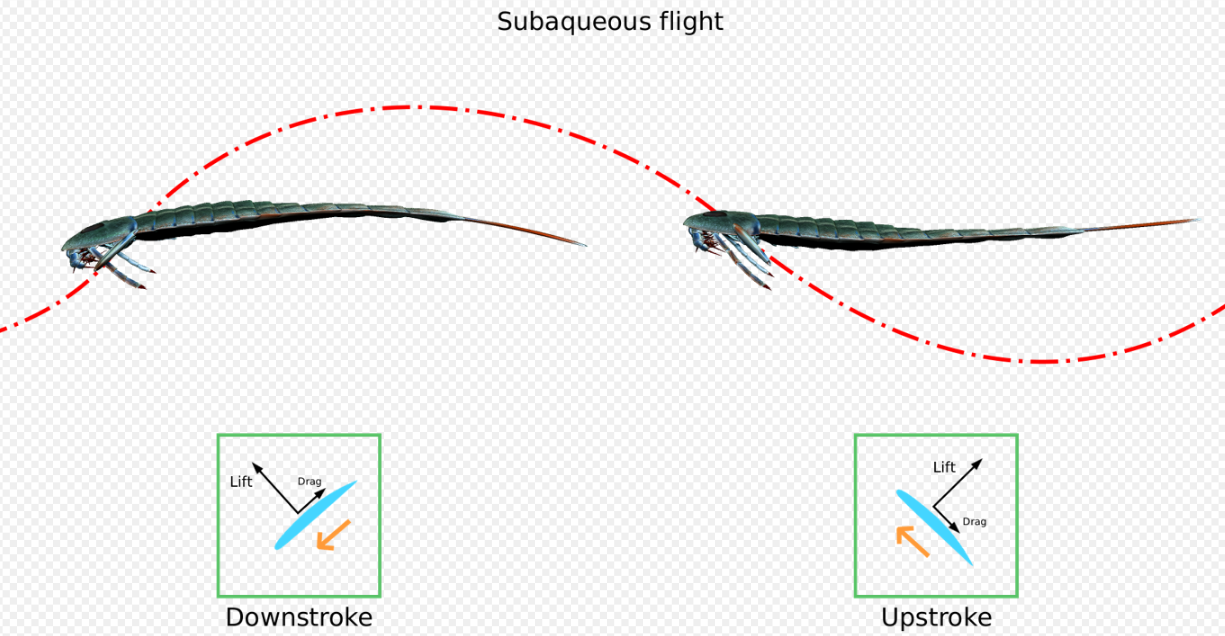
An alternative hypothesis for Eurypterus swimming behavior is that individuals were capable of underwater flying (or subaqueous flight), in which the sinuous motions and shape of the paddles themselves acting as hydrofoils are enough to generate lift.[28] This type is similar to that found in sea turtles and sea lions. It has a relatively slower acceleration rate than the rowing type, especially since adults have proportionally smaller paddles than juveniles. But since the larger sizes of adults mean a higher drag coefficient, using this type of propulsion is more energy-efficient.[22][29]
Juveniles probably swam using the rowing type, the rapid acceleration afforded by this propulsion is more suited for quickly escaping predators. A small 16.5 cm (6.5 in) Eurypterus could achieve two and a half body lengths per second immediately.[22] Larger adults, meanwhile, probably swam with the subaqueous flight type. The adults slightly faster than turtles and sea otters.[29][30]
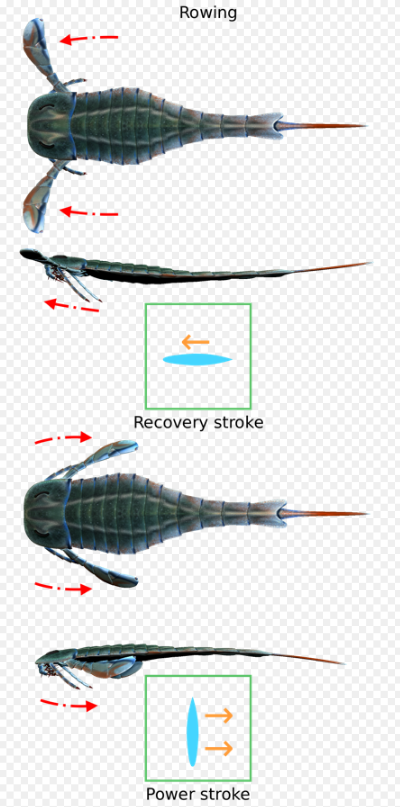
Trace fossil evidence indicates that Eurypterus employed a rowing stroke when in close proximity to the seafloor.[31] Arcuites bertiensis is an ichnospecies that includes a pair of crescent-shaped impressions and a short medial drag, and it has been found in upper Silurian eurypterid Lagerstatten in Ontario and Pennsylvania. This trace fossil is very similar to traces made by modern aquatic swimming insects that row such as water boatmen, and is considered to have been made by juvenile to adult-sized eurypterids while swimming in very shallow nearshore marine environments. The morphology of A. bertiensis suggests that Eurypterus had the ability to move its swimming appendages in both the horizontal and vertical plane.
Eurypterus did not swim to hunt, rather they simply swam in order to move from one feeding site to another quickly. Most of the time they walked on the substrate with their legs (including their swimming leg). They were generalist species, equally likely to engage in predation or scavenging. They hunted small soft-bodied invertebrates like worms. They utilized the mass of spines on their front appendages to both kill and hold them while they used their chelicerae to rip off pieces small enough to swallow. Young individuals may also have fallen prey to cannibalism by larger adults.[22]
Eurypterus were most probably marine animals, as their remains are mostly found in intertidal shallow environments. The concentrations of Eurypterus fossils in certain sites has been interpreted to be a result of mass mating and molting behavior. Juveniles were likely to have inhabited nearshore hypersaline environments, safer from predators, and moved to deeper waters as they grew older and larger. Adults that reach sexual maturity would then migrate en masse to shore areas in order to mate, lay eggs, and molt. Activities that would have made them more vulnerable to predators. This could also explain why the vast majority of fossils found in such sites are molts and not of actual animals. The same behavior can be seen in modern horseshoe crabs.[11]
5.1. Respiration

Examinations of the respiratory systems of Eurypterus have led many paleontologists to conclude that it was capable of breathing air and walking on land for a short amount of time. Eurypterus had two types of respiratory systems. Its main organs for breathing were the book gills inside the segments of the mesosoma. These structures were supported by semicircular 'ribs' and were probably attached near the center of the body, similar to the gills of modern horseshoe crabs.[19] They were protected under platelike appendages (which actually formed the apparent 'belly' of Eurypterus) known as Blatfüsse.[32] These gills may have also played a role in osmoregulation.[22]
The second system are the Kiemenplatten, also referred to as gill-tracts. These oval-shaped areas within the body wall of the preabdomen. Their surfaces are covered with numerous small spines arranged into hexagonal 'rosettes'. These areas were vascularized, hence the conclusion that they were secondary breathing organs.[20]
The function of the book gills are usually interpreted to be for aquatic breathing, while the Kiemenplatten are supplementary for temporary breathing on land.[19] However, some authors have argued that the two systems alone could not have supported an organism the size of Eurypterus. Both structures might actually have been for breathing air and the true gills (for underwater breathing) of Eurypterus have yet to be discovered.[20][32] Eurypterus, however, were undoubtedly primarily aquatic.[32]
5.2. Ontogeny
Juvenile Eurypterus differed from adults in several ways. Their carapaces were narrower and longer (parabolic) in contrast to the trapezoidal carapaces of adults. The eyes are aligned almost laterally but move to a more anterior location during growth. The preabdomen also lengthened, increasing the overall length of the ophisthosoma. The swimming legs also became narrower and the telsons shorter and broader (though in E. tetragonophthalmus and E. henningsmoeni the telsons changed from being angular in juveniles to larger and more rounded in adults). All these changes are believed to be a result of the respiratory and reproductive requirements of adults.[23]
6. Paleoecology

Members of Eurypterus existed for a relatively short time, yet they are the most abundant eurypterids found today.[5] They flourished between the Late Llandovery epoch (around 432 million years ago) to sometime during the Přídolí epoch (418.1 million years ago) of the Silurian period. A span of only around 10 to 14 million years.[23][33]
During this period, the landmasses were mostly restricted to the southern hemisphere of the Earth, with the supercontinent Gondwana straddling the South Pole. The equator had three continents (Avalonia, Baltica, and Laurentia) which slowly drifted together to form the second supercontinent of Laurussia (also known as Euramerica, not to be confused with Laurasia).[2]
The ancestors of Eurypterus were believed to have originated from Baltica (eastern Laurussia, modern western Eurasia) based on the earliest recorded fossils. During the Silurian, they spread to Laurentia (western Laurussia, modern North America) when the two continents began to collide. They rapidly colonized the continent as invasive species, becoming the most dominant eurypterid in the region. This accounts for why they are the most commonly found genus of eurypterids today. Eurypterus (and other members of Eurypteroidea), however, were unable to cross vast expanses of oceans between the two supercontinents during the Silurian. Their range were thus limited to the coastlines and the large, shallow, and hypersaline inland seas of Laurussia.[2][5]
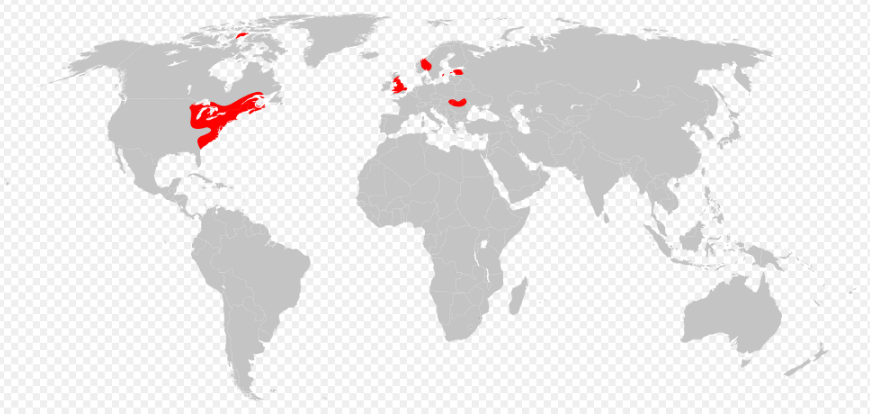
They are now only known from fossils from North America, Europe, and northwestern Asia, cratons that were the former components of Laurussia. While three species of Eurypterus were purportedly discovered in China in 1957, the evidence of them belonging to the genus (or if they were even eurypterids at all) is nonexistent. No other traces of Eurypterus in modern continents from Gondwana are currently known.[5]
Eurypterus are very common fossils in their regions of occurrence, millions of specimens are possible in a given area, though access to the rock formations may be difficult.[34] Most fossil eurypterids are the disjointed shed exoskeleton (known as exuviae) of individuals after molting (ecdysis). Some are complete but are most probably exuviae as well. Fossils of the actual remains of eurypterids (i.e. their carcasses) are relatively rare.[2] Fossil eurypterids are often deposited in characteristic windrows, probably a result of wave and wind action.[35]
The content is sourced from: https://handwiki.org/wiki/Biology:Eurypterus
References
- John R. Nudds; Paul Selden (2008). Fossil ecosystems of North America: a guide to the sites and their extraordinary biotas. Manson Publishing. pp. 74, 78–82. ISBN 978-1-84076-088-0. https://books.google.com/books?id=_z-ca7JAVJIC&pg=PA78.
- O.E. Tetlie; I. Rábano (2007). "Specimens of Eurypterus (Chelicerata, Eurypterida) in the collections of Museo Geominero (Geological Survey of Spain), Madrid". Boletín Geológico y Minero 118 (1): 117–126. ISSN 0366-0176. http://www.igme.es/internet/Boletin/2007/118_1_2007/ART.%209.pdf. Retrieved May 22, 2011.
- "Eurypterida: Fossil Record". University of California Museum of Paleontology. http://www.ucmp.berkeley.edu/arthropoda/chelicerata/eurypteridfr.html.
- Erik N. Kjellesvig-Waering (1958). "The genera, species and subspecies of the family Eurypteridae, Burmeister, 1845". Journal of Paleontology 32 (6): 1107–1148. ISSN 0022-3360. http://www.worldcat.org/issn/0022-3360
- O. Erik Tetlie (2007). "Distribution and dispersal history of Eurypterida (Chelicerata)". Palaeogeography, Palaeoclimatology, Palaeoecology 252 (3–4): 557–574. doi:10.1016/j.palaeo.2007.05.011. ISSN 0031-0182. http://fossilinsects.net/pdfs/tetlie_2007_PX3_DistribDispersalEurypterida.pdf. Retrieved May 20, 2011.
- "New York State Fossil – Eurypterus remipes". New York State Library. April 27, 2009. http://www.nysl.nysed.gov/emblems/fossil.htm.
- Daniel Cressey (2007). "Giant sea scorpion discovered". Nature. doi:10.1038/news.2007.272. http://www.nature.com/news/2007/071120/full/news.2007.272.html. Retrieved May 22, 2011.
- Samuel J. Ciurca, Jr.. "The size that common Eurypterus species attained (Late Silurian Bertie Group of New York and Ontario, Canada)". Eurypterids.net. http://eurypterids.net/EurypteridMonth5.html.
- Ruebenstahl, Alexander; Ciurca, Jr., Samuel J.; Briggs, Derek E. G. (2021). "A giant Eurypterus from the Silurian (Pridoli) Bertie Group of North America". Bulletin of the Peabody Museum of Natural History 62 (1): 3–13. doi:10.3374/014.062.0101. https://bioone.org/journals/bulletin-of-the-peabody-museum-of-natural-history/volume-62/issue-1/014.062.0101/A-Giant-Eurypterus-from-the-Silurian-Pridoli-Bertie-Group-of/10.3374/014.062.0101.short.
- "Eurypterus remipes NPL4415". Texas Natural Science Center: Non-vertebrate Paleontology, The University of Texas. http://www.utexas.edu/tmm/npl/exhibits/eurypterus.html.
- Conrad Burkert. Environment preference of eurypterids – indications for freshwater adaptation?. Technische Universität Bergakademie Freiberg. http://www.geo.tu-freiberg.de/oberseminar/os06_07/Conrad_Burkert.pdf. Retrieved May 21, 2011.
- Pat Vickers Rich, Mildred Adams Fenton, Carroll Lane Fenton, & Thomas Hewitt Rich (1989). The fossil book: a record of prehistoric life. Courier Dover Publications. p. 225. ISBN 978-0-486-29371-4. https://books.google.com/books?id=_ntSspji0LYC&pg=PA225.
- James Lamsdell. "Eurypterid Basics". Eurypterids.co.uk. http://www.eurypterids.co.uk/eurypterids.htm.
- V. P. Tollerton, Jr. (1989). "Morphology, Taxonomy, and Classification of the Order Eurypterida Burmeister, 1843". Journal of Paleontology 63 (5): 642–657. doi:10.1017/S0022336000041275. ISSN 0022-3360. https://dx.doi.org/10.1017%2FS0022336000041275
- "Eurypterid". The Encyclopedia Americana: Egypt to Falsetto. 10. Grolier. 1983. pp. 708–709. ISBN 978-0-7172-0114-3.
- "Les scorpions de mer (les Eurypterida)" (in fr). Paleopedia. http://paleopedia.free.fr/invert%E9br%E9_arthropode_eurypteridae.html.
- Roy Plotnick. Eurypterida. Access Science. doi:10.1036/1097-8542.246600. http://accessscience.com/content/Eurypterida/246600. Retrieved May 23, 2011.
- Jason A. Dunlop; Mark Webster (1999). "Fossil Evidence, Terrestrialization and Arachnid Phylogeny". The Journal of Arachnology 27: 86–93. ISSN 0161-8202. http://www.museunacional.ufrj.br/mndi/Aracnologia/pdfliteratura/Dunlop%20&%20Webster%20JOA%201999%20Fossil%20Arach%20terrestrialization.pdf. Retrieved May 23, 2011.
- Phillip L. Manning; Jason A. Dunlop (1995). "The respiratory organs of Eurypterids". Palaeontology 38 (2): 287–297. ISSN 0031-0239. http://palaeontology.palass-pubs.org/pdf/Vol%2038/Pages%20287-297.pdf. Retrieved May 21, 2011.
- P. A. Selden (1985). "Eurypterid respiration". Philosophical Transactions of the Royal Society of London B 309 (1138): 219–226. doi:10.1098/rstb.1985.0081. ISSN 0080-4622. Bibcode: 1985RSPTB.309..219S. http://homepage.mac.com/paulselden/Sites/Website/EuryResp.pdf. Retrieved May 22, 2011.
- Lief Størmer (1955). "Merostomata". in C. Raymond. Treatise on Invertebrate Paleontology, Part P: Arthropoda 2: Chelicerata, Pycnogonida & Palaeoisopus. Geological Society of America and University of Kansas Press. pp. 31–34. ISBN 978-0-8137-3016-5.
- Selden, Paul A. (1981). "Functional morphology of the prosoma of Baltoeurypterus tetragonophthalmus (Fischer) (Chelicerata: Eurypterida)". Transactions of the Royal Society of Edinburgh: Earth Sciences 72: 9–48. doi:10.1017/s0263593300003217. https://www.cambridge.org/core/journals/earth-and-environmental-science-transactions-of-royal-society-of-edinburgh/article/functional-morphology-of-the-prosoma-of-baltoeurypterus-tetragonophthalmus-fischer-chelicerata-eurypterida/D47ECC0D4657530A7931691566F08D5F. Retrieved February 9, 2018.
- O. Erik Tetlie (2006). "Two new Silurian species of Eurypterus (Chelicerata: Eurypterida) from Norway and Canada and the phylogeny of the genus". Journal of Systematic Palaeontology 4 (4): 397–412. doi:10.1017/S1477201906001921. ISSN 1478-0941. http://www.museunacional.ufrj.br/mndi/Aracnologia/Eurypterida/Tetlie%202006%20Silurian%20Eurypterus.pdf. Retrieved May 20, 2011.
- Jason A. Dunlop, David Penney, & Denise Jekel; with additional contributions from Lyall I. Anderson, Simon J. Braddy, James C. Lamsdell, Paul A. Selden, & O. Erik Tetlie (2011). "A summary list of fossil spiders and their relatives". in Norman I. Platnick. The world spider catalog, version 11.5. American Museum of Natural History. http://research.amnh.org/iz/spiders/catalog/Fossil11.5.pdf. Retrieved May 21, 2011.
- P. Weygoldt; H. F. Paulus (2009). "Untersuchungen zur Morphologie, Taxonomie und Phylogenie der Chelicerata† II. Cladogramme und die Entfaltung der Chelicerata". Journal of Zoological Systematics and Evolutionary Research 17 (3): 177–200. doi:10.1111/j.1439-0469.1979.tb00699.x. ISSN 1439-0469. https://dx.doi.org/10.1111%2Fj.1439-0469.1979.tb00699.x
- "Eurypterida: Systematics". University of California Museum of Paleontology. http://www.ucmp.berkeley.edu/arthropoda/chelicerata/eurypteridsy.html.
- John W. Merck, Jr.. "The biomechanics of swimming". Department of Geology, University of Maryland. http://www.geol.umd.edu/~jmerck/bsci392/lecture10/lecture10.html.
- Plotnick, Roy E. (1985). "Lift based mechanisms for swimming in eurypterids and portunid crabs" (in en). Earth and Environmental Science Transactions of the Royal Society of Edinburgh 76 (2–3): 325–337. doi:10.1017/S0263593300010543. ISSN 1755-6929. https://www.cambridge.org/core/journals/earth-and-environmental-science-transactions-of-royal-society-of-edinburgh/article/lift-based-mechanisms-for-swimming-in-eurypterids-and-portunid-crabs/89954954F4CB25CD197F5BC4BB1F55D6.
- John W. Merck, Jr.. "Eurypterids, arachnids, and the arthropod invasion of the land". Department of Geology, University of Maryland. http://www.geol.umd.edu/~jmerck/bsci392/lecture25/lecture25.html.
- "The story of one man's plight to make fossils swim". http://gknight.tripod.com/.
- Vrazo, Matthew B.; Ciurca, Jr., Samuel J. (2018-03-01). "New trace fossil evidence for eurypterid swimming behaviour" (in en). Palaeontology 61 (2): 235–252. doi:10.1111/pala.12336. ISSN 1475-4983. https://dx.doi.org/10.1111%2Fpala.12336
- Paul A. Selden; Andrew J. Jeram (1989). "Palaeophysiology of terrestrialisation in the Chelicerata". Transactions of the Royal Society of Edinburgh: Earth Sciences 80 (3–4): 303–310. doi:10.1017/s0263593300028741. ISSN 0263-5933. http://homepage.mac.com/paulselden/Sites/Website/Palaeophysiology.pdf. Retrieved May 23, 2011.
- "Eurypterus". Paleobiology Database. http://paleodb.org/cgi-bin/bridge.pl?a=checkTaxonInfo&taxon_no=18968&is_real_user=1.
- Samuel J. Ciurca, Jr.. "Eurypterus lacustris Fauna". Eurypterids.net. http://eurypterids.net/EurypteruslacustrisFauna.html.
- Samuel J. Ciurca, Jr.. "Commentary on a specimen of an Eurypterid". Eurypterids.net. http://eurypterids.net/EurypterusLacustris4.html.