Chloride intracellular channel proteins (CLICs are the dimorphic protein present in both soluble and membrane fractions. As an integral membrane protein, CLICs potentially possess ion channel activity. In vertebrates, CLICs are classified into six classes: CLIC1, 2, 3, 4, 5, and 6. CLIC2 is expressed at higher levels in benign tumors than in malignant ones, most likely preventing tumor cell invasion into surrounding tissues. CLIC2 is also expressed in the vascular endothelial cells of normal tissues and maintains their intercellular adhesive junctions, presumably suppressing the hematogenous metastasis of malignant tumor cells.
- metastasis
- invasion
- MMP
- MT1-MMP
- CLIC4
- glioma
1. Introduction
1.1. CLIC Family
| CLICs | Distribution | Ion Channel | Biological Function | References |
|---|---|---|---|---|
| CLIC1 | Various organs | Poorly selective anion channels | Participates in inflammatory processes Activation of MYC signaling Enzymatic activity Redox regulation |
[11][12][13][14][15][16][17] |
| CLIC2 | Blood vessels. heart, liver | Anion channels | Modulation of ryanodine receptor Inhibition of MMP14 activity |
[4][5][13][18][19] |
| CLIC3 | Muscles, heart, lung, kidney | Component of anion channel, regulator of channel | Endosomal trafficking Promote invasive behavior |
[20][21][22][23] |
| CLIC4 | Various organs | Poorly selective ion channels | Enhance tumor invasiveness Enhance TGF-β signaling Induce apoptosis Involved in angiogenesis Stimulation of MMP14 activity |
[24][25][26][27][28][29] |
| CLIC5 | Kidney, heart, lung, colon | Poorly selective ion channels | Actin cytoskeleton-dependent membrane remodeling | [9][25][30] |
| CLIC6 | Soluble and membrane fractions | Unknown | Interact with dopamine receptors | [31][32][33] |
1.2. Known Structures of CLIC2
1.3. Known Functions of CLIC2
1.4. CLIC2 and Malignancy
| Cancer | Breast | Ovarian | Lung | Gastric | Liver | Pancreatic |
|---|---|---|---|---|---|---|
| CLIC1 | detrimental | detrimental | n.s. | ameliorative | detrimental | detrimental |
| CLIC2 | ameliorative | detrimental | ameliorative | ameliorative | ameliorative | n.s. |
| CLIC3 | detrimental | detrimental | n.s. | detrimental | n.s. | detrimental |
| CLIC4 | n.s. | detrimental | detrimental | detrimental | n.s. | detrimental |
| CLIC5 | ameliorative | detrimental | ameliorative | ameliorative | n.s. | n.s. |
| CLIC6 | ameliorative | ameliorative | ameliorative | ameliorative | n.s. | n.s. |
1.5. Why CLIC2?
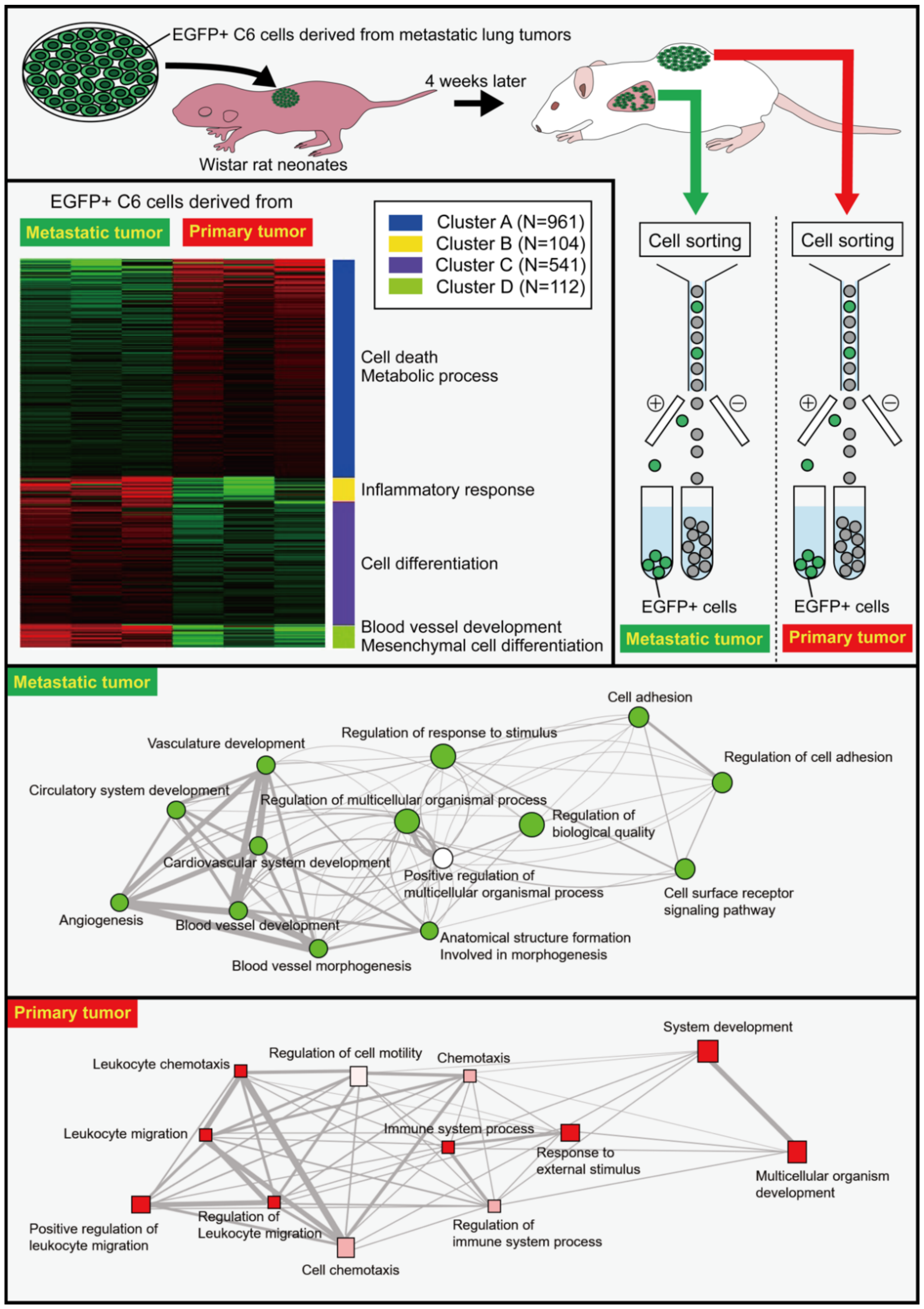
2. Roles of Secreted CLIC2; Relationship with MMPs
3. Intercellular Adhesive Structures and CLIC2
4. Modulation of MMP Activities by CLIC4
This entry is adapted from the peer-reviewed paper 10.3390/cancers14194890
References
- Littler, D.R.; Harrop, S.J.; Goodchild, S.C.; Phang, J.M.; Mynott, A.V.; Jiang, L.; Valenzuela, S.M.; Mazzanti, M.; Brown, L.J.; Breit, S.N.; et al. The enigma of the CLIC proteins: Ion channels, redox proteins, enzymes, scaffolding proteins? FEBS Lett. 2010, 584, 2093–2101.
- Zeng, J.; Li, Z.; Lui, E.Y.; Lam, S.H.; Swaminathan, K. Tilapia and human CLIC2 structures are highly conserved. Biochem. Biophys. Res. Commun. 2018, 495, 1752–1757.
- Argenzio, E.; Moolenaar, W.H. Emerging biological roles of Cl- intracellular channel proteins. J. Cell Sci. 2016, 129, 4165–4174.
- Ozaki, S.; Umakoshi, A.; Yano, H.; Ohsumi, S.; Sumida, Y.; Hayase, E.; Usa, E.; Islam, A.; Choudhury, M.E.; Nishi, Y.; et al. Chloride intracellular channel protein 2 is secreted and inhibits MMP14 activity, while preventing tumor cell invasion and metastasis. Neoplasia 2021, 23, 754–765.
- Ueno, Y.; Ozaki, S.; Umakoshi, A.; Yano, H.; Choudhury, M.E.; Abe, N.; Sumida, Y.; Kuwabara, J.; Uchida, R.; Islam, A.; et al. Chloride intracellular channel protein 2 in cancer and non-cancer human tissues: Relationship with tight junctions. Tissue Barriers 2019, 7.
- Ulmasov, B.; Bruno, J.; Gordon, N.; Hartnett, M.E.; Edwards, J.C. Chloride intracellular channel protein-4 functions in angiogenesis by supporting acidification of vacuoles along the intracellular tubulogenic pathway. Am. J. Pathol. 2009, 174, 1084–1096.
- Berry, K.L.; Bulow, H.E.; Hall, D.H.; Hobert, O. A C. elegans CLIC-like protein required for intracellular tube formation and maintenance. Science 2003, 302, 2134–2137.
- Fernandez-Salas, E.; Sagar, M.; Cheng, C.; Yuspa, S.H.; Weinberg, W.C. p53 and tumor necrosis factor alpha regulate the expression of a mitochondrial chloride channel protein. J. Biol. Chem. 1999, 274, 36488–36497.
- Berryman, M.; Bretscher, A. Identification of a novel member of the chloride intracellular channel gene family (CLIC5) that associates with the actin cytoskeleton of placental microvilli. Mol. Biol. Cell 2000, 11, 1509–1521.
- Tang, T.; Lang, X.; Xu, C.; Wang, X.; Gong, T.; Yang, Y.; Cui, J.; Bai, L.; Wang, J.; Jiang, W.; et al. CLICs-dependent chloride efflux is an essential and proximal upstream event for NLRP3 inflammasome activation. Nat. Commun. 2017, 8, 202.
- Singh, H.; Ashley, R.H. Redox regulation of CLIC1 by cysteine residues associated with the putative channel pore. Biophys. J. 2006, 90, 1628–1638.
- Tulk, B.M.; Kapadia, S.; Edwards, J.C. CLIC1 inserts from the aqueous phase into phospholipid membranes, where it functions as an anion channel. Am. J. Physiol. Cell Physiol. 2022, 282, C1103–C1112.
- Littler, D.R.; Harrop, S.J.; Fairlie, W.D.; Brown, L.J.; Pankhurst, G.J.; Pankhurst, S.; DeMaere, M.Z.; Campbell, T.J.; Bauskin, A.R.; Tonini, R.; et al. The intracellular chloride ion channel protein CLIC1 undergoes a redox-controlled structural transition. J. Biol. Chem. 2004, 279, 9298–9305.
- Peter, B.; Fanucchi, S.; Dirr, H.W. A conserved cationic motif enhances membrane binding and insertion of the chloride intracellular channel protein 1 transmembrane domain. Eur. Biophys. J. 2014, 43, 405–414.
- Jiang, L.; Salao, K.; Li, H.; Rybicka, J.M.; Yates, R.M.; Luo, X.W.; Shi, X.X.; Kuffner, T.; Tsai, V.W.; Husaini, Y.; et al. Intracellular chloride channel protein CLIC1 regulates macrophage function through modulation of phagosomal acidification. J. Cell Sci. 2012, 125 Pt 22, 5479–5488.
- Salao, K.; Jiang, L.; Li, H.; Tsai, V.W.; Husaini, Y.; Curmi, P.M.; Brown, L.J.; Brown, D.A.; Breit, S.N. CLIC1 regulates dendritic cell antigen processing and presentation by modulating phagosome acidification and proteolysis. Biol. Open 2016, 5, 620–630.
- Al Khamici, H.; Brown, L.J.; Hossain, K.R.; Hudson, A.L.; Sinclair-Burton, A.A.; Ng, J.P.; Daniel, E.L.; Hare, J.E.; Cornell, B.A.; Curmi, P.M.; et al. Members of the chloride intracellular ion channel protein family demonstrate glutaredoxin-like enzymatic activity. PLoS ONE 2015, 10, e115699.
- Board, P.G.; Coggan, M.; Watson, S.; Gage, P.W.; Dulhunty, A.F. CLIC-2 modulates cardiac ryanodine receptor Ca2+ release channels. Int. J. Biochem. Cell Biol. 2004, 36 Pt 18, 1599–1612.
- Cromer, B.A.; Gorman, M.A.; Hansen, G.; Adams, J.J.; Coggan, M.; Littler, D.R.; Brown, L.J.; Mazzanti, M.; Breit, S.N.; Curmi, P.M.; et al. Structure of the Janus protein human CLIC2. J. Mol. Biol. 2007, 374, 719–731.
- Qian, Z.; Okuhara, D.; Abe, M.K.; Rosner, M.R. Molecular cloning and characterization of a mitogen-activated protein kinase-associated intracellular chloride channel. J. Biol. Chem. 1999, 274, 1621–1627.
- Dozynkiewicz, M.A.; Jamieson, N.B.; Macpherson, I.; Grindlay, J.; van den Berghe, P.V.; von Thun, A.; Morton, J.P.; Gourley, C.; Timpson, P.; Nixon, C.; et al. Rab25 and CLIC3 collaborate to promote integrin recycling from late endosomes/lysosomes and drive cancer progression. Dev. Cell 2012, 22, 131–145.
- Macpherson, I.R.; Rainero, E.; Mitchell, L.E.; van den Berghe, P.V.; Speirs, C.; Dozynkiewicz, M.A.; Chaudhary, S.; Kalna, G.; Edwards, J.; Timpson, P.; et al. CLIC3 controls recycling of late endosomal MT1-MMP and dictates invasion and metastasis in breast cancer. J. Cell Sci. 2014, 127, 3893–3901.
- Hernandez-Fernaud, J.R.; Ruengeler, E.; Casazza, A.; Neilson, L.J.; Pulleine, E.; Santi, A.; Ismail, S.; Lilla, S.; Dhayade, S.; MacPherson, I.R.; et al. Secreted CLIC3 drives cancer progression through its glutathione-dependent oxidoreductase activity. Nat. Commun. 2017, 8, 14206.
- Duncan, R.R.; Westwood, P.K.; Boyd, A.; Ashley, R.H. Rat brain p64H1, expression of a new member of the p64 chloride channel protein family in endoplasmic reticulum. J. Biol. Chem. 1997, 272, 23880–23886.
- Singh, H.; Ashley, R.H. CLIC4 (p64H1) and its putative transmembrane domain form poorly selective, redox-regulated ion channels. Mol. Membr. Biol. 2007, 24, 41–52.
- Shukla, A.; Edwards, R.; Yang, Y.; Hahn, A.; Folkers, K.; Ding, J.; Padmakumar, V.C.; Cataisson, C.; Suh, K.S.; Yuspa, S.H. CLIC4 regulates TGF-beta-dependent myofibroblast differentiation to produce a cancer stroma. Oncogene 2014, 33, 842–850.
- Fernandez-Salas, E.; Suh, K.S.; Speransky, V.V.; Bowers, W.L.; Levy, J.M.; Adams, T.; Pathak, K.R.; Edwards, L.E.; Hayes, D.D.; Cheng, C.; et al. mtCLIC/CLIC4, an organellular chloride channel protein, is increased by DNA damage and participates in the apoptotic response to p53. Mol. Cell. Biol. 2002, 22, 3610–3620.
- Tung, J.J.; Hobert, O.; Berryman, M.; Kitajewski, J. Chloride intracellular channel 4 is involved in endothelial proliferation and morphogenesis in vitro. Angiogenesis 2009, 12, 209–220.
- Hsu, K.S.; Otsu, W.; Li, Y.; Wang, H.C.; Chen, S.; Tsang, S.H.; Chuang, J.Z.; Sung, C.H. CLIC4 regulates late endosomal trafficking and matrix degradation activity of MMP14 at focal adhesions in RPE cells. Sci Rep. 2019, 9, 12247.
- Salles, F.T.; Andrade, L.R.; Tanda, S.; Grati, M.; Plona, K.L.; Gagnon, L.H.; Johnson, K.R.; Kachar, B.; Berryman, M.A. CLIC5 stabilizes membrane-actin filament linkages at the base of hair cell stereocilia in a molecular complex with radixin, taperin, and myosin VI. Cytoskeleton 2014, 71, 61–78.
- Ferofontov, A.; Strulovich, R.; Marom, M.; Giladi, M.; Haitin, Y. Inherent flexibility of CLIC6 revealed by crystallographic and solution studies. Sci. Rep. 2018, 8, 6882.
- Griffon, N.; Jeanneteau, F.; Prieur, F.; Diaz, J.; Sokoloff, P. CLIC6, a member of the intracellular chloride channel family, interacts with dopamine D(2)-like receptors. Mol. Brain Res. 2003, 117, 47–57.
- Friedli, M.; Guipponi, M.; Bertrand, S.; Bertrand, D.; Neerman-Arbez, M.; Scott, H.S.; Antonarakis, S.E.; Reymond, A. Identification of a novel member of the CLIC family, CLIC6, mapping to 21q22.12. Gene 2003, 320, 31–40.
- Littler, D.R.; Assaad, N.N.; Harrop, S.J.; Brown, L.J.; Pankhurst, G.J.; Luciani, P.; Aguilar, M.I.; Mazzanti, M.; Berryman, M.A.; Breit, S.N.; et al. Crystal structure of the soluble form of the redox-regulated chloride ion channel protein CLIC4. FEBS J. 2005, 272, 4996–5007.
- Jalilian, C.; Gallant, E.M.; Board, P.G.; Dulhunty, A.F. Redox potential and the response of cardiac ryanodine receptors to CLIC-2, a member of the glutathione S-transferase structural family. Antioxid. Redox. Signal. 2008, 10, 1675–1686.
- Dulhunty, A.; Gage, P.; Curtis, S.; Chelvanayagam, G.; Board, P. The glutathione transferase structural family includes a nuclear chloride channel and a ryanodine receptor calcium release channel modulator. J. Biol. Chem. 2001, 276, 3319–3323.
- Molina-Navarro, M.M.; Rosello-Lleti, E.; Ortega, A.; Tarazon, E.; Otero, M.; Martinez-Dolz, L.; Lago, F.; Gonzalez-Juanatey, J.R.; Espana, F.; Garcia-Pavia, P.; et al. Differential gene expression of cardiac ion channels in human dilated cardiomyopathy. PLoS ONE 2013, 8, e79792.
- Pierchala, B.A.; Munoz, M.R.; Tsui, C.C. Proteomic analysis of the slit diaphragm complex: CLIC5 is a protein critical for podocyte morphology and function. Kidney Int. 2010, 78, 868–882.
- Rickhag, M.; Wieloch, T.; Gido, G.; Elmer, E.; Krogh, M.; Murray, J.; Lohr, S.; Bitter, H.; Chin, D.J.; von Schack, D.; et al. Comprehensive regional and temporal gene expression profiling of the rat brain during the first 24 h after experimental stroke identifies dynamic ischemia-induced gene expression patterns, and reveals a biphasic activation of genes in surviving tissue. J. Neurochem. 2006, 96, 14–29.
- Suh, K.S.; Mutoh, M.; Mutoh, T.; Li, L.; Ryscavage, A.; Crutchley, J.M.; Dumont, R.A.; Cheng, C.; Yuspa, S.H. CLIC4 mediates and is required for Ca2+-induced keratinocyte differentiation. J. Cell Sci. 2007, 120 Pt 15, 2631–2640.
- Shukla, A.; Malik, M.; Cataisson, C.; Ho, Y.; Friesen, T.; Suh, K.S.; Yuspa, S.H. TGF-beta signalling is regulated by Schnurri-2-dependent nuclear translocation of CLIC4 and consequent stabilization of phospho-Smad2 and 3. Nat. Cell Biol. 2009, 11, 777–784.
- Gururaja Rao, S.; Patel, N.J.; Singh, H. Intracellular Chloride Channels: Novel Biomarkers in Diseases. Front. Physiol. 2020, 11, 96.
- Ponnalagu, D.; Rao, S.G.; Farber, J.; Xin, W.; Hussain, A.T.; Shah, K.; Tanda, S.; Berryman, M.A.; Edwards, J.C.; Singh, H. Data supporting characterization of CLIC1, CLIC4, CLIC5 and DmCLIC antibodies and localization of CLICs in endoplasmic reticulum of cardiomyocytes. Data Brief. 2016, 7, 1038–1044.
- Edwards, J.C.; Kahl, C.R. Chloride channels of intracellular membranes. FEBS Lett. 2010, 584, 2102–2111.
- Gururaja Rao, S.; Ponnalagu, D.; Patel, N.J.; Singh, H. Three Decades of Chloride Intracellular Channel Proteins: From Organelle to Organ Physiology. Curr. Protoc. Pharmacol. 2018, 80, 11–21.
- Singh, H. Two decades with dimorphic Chloride Intracellular Channels (CLICs). FEBS Lett. 2010, 584, 2112–2121.
- Kim, H.J.; Lee, P.C.; Hong, J.H. Chloride Channels and Transporters: Roles beyond Classical Cellular Homeostatic pH or Ion Balance in Cancers. Cancers 2022, 14, 856.
- Takano, K.; Liu, D.; Tarpey, P.; Gallant, E.; Lam, A.; Witham, S.; Alexov, E.; Chaubey, A.; Stevenson, R.E.; Schwartz, C.E.; et al. An X-linked channelopathy with cardiomegaly due to a CLIC2 mutation enhancing ryanodine receptor channel activity. Hum. Mol. Genet. 2012, 21, 4497–4507.
- Witham, S.; Takano, K.; Schwartz, C.; Alexov, E. A missense mutation in CLIC2 associated with intellectual disability is predicted by in silico modeling to affect protein stability and dynamics. Proteins 2011, 79, 2444–2454.
- Richardson, S.J.; Steele, G.A.; Gallant, E.M.; Lam, A.; Schwartz, C.E.; Board, P.G.; Casarotto, M.G.; Beard, N.A.; Dulhunty, A.F. Association of FK506 binding proteins with RyR channels-effect of CLIC2 binding on sub-conductance opening and FKBP binding. J. Cell Sci. 2017, 130, 3588–3600.
- Meng, X.; Wang, G.; Viero, C.; Wang, Q.; Mi, W.; Su, X.D.; Wagenknecht, T.; Williams, A.J.; Liu, Z.; Yin, C.C. CLIC2-RyR1 interaction and structural characterization by cryo-electron microscopy. J. Mol. Biol. 2009, 387, 320–334.
- Suh, K.S.; Mutoh, M.; Gerdes, M.; Crutchley, J.M.; Mutoh, T.; Edwards, L.E.; Dumont, R.A.; Sodha, P.; Cheng, C.; Glick, A.; et al. Antisense suppression of the chloride intracellular channel family induces apoptosis, enhances tumor necrosis factor α-induced apoptosis, and inhibits tumor growth. Cancer Res. 2005, 65, 562–571.
- Kuwabara, J.; Umakoshi, A.; Abe, N.; Sumida, Y.; Ohsumi, S.; Usa, E.; Taguchi, K.; Choudhury, M.E.; Yano, H.; Matsumoto, S.; et al. Truncated CD200 stimulates tumor immunity leading to fewer lung metastases in a novel Wistar rat metastasis model. Biochem. Biophys. Res. Commun. 2018, 496, 542–548.
- Xu, T.; Wang, Z.; Dong, M.; Wu, D.; Liao, S.; Li, X. Chloride intracellular channel protein 2: Prognostic marker and correlation with PD-1/PD-L1 in breast cancer. Aging 2020, 12, 17305–17327.
- Liu, J.; Jin, X.; Liu, K.J.; Liu, W. Matrix metalloproteinase-2-mediated occludin degradation and caveolin-1-mediated claudin-5 redistribution contribute to blood-brain barrier damage in early ischemic stroke stage. J. Neurosci. 2012, 32, 3044–3057.
- Romanic, A.M.; White, R.F.; Arleth, A.J.; Ohlstein, E.H.; Barone, F.C. Matrix metalloproteinase expression increases after cerebral focal ischemia in rats: Inhibition of matrix metalloproteinase-9 reduces infarct size. Stroke 1998, 29, 1020–1030.
- Yang, Y.; Estrada, E.Y.; Thompson, J.F.; Liu, W.; Rosenberg, G.A. Matrix metalloproteinase-mediated disruption of tight junction proteins in cerebral vessels is reversed by synthetic matrix metalloproteinase inhibitor in focal ischemia in rat. J. Cereb. Blood Flow Metab. 2007, 27, 697–709.
- Miyamori, H.; Takino, T.; Kobayashi, Y.; Tokai, H.; Itoh, Y.; Seiki, M.; Sato, H. Claudin promotes activation of pro-matrix metalloproteinase-2 mediated by membrane-type matrix metalloproteinases. J. Biol. Chem. 2001, 276, 28204–28211.
- Mao, Y.; Kleinjan, M.L.; Jilishitz, I.; Swaminathan, B.; Obinata, H.; Komarova, Y.A.; Bayless, K.J.; Hla, T.; Kitajewski, J.K. CLIC1 and CLIC4 mediate endothelial S1P receptor signaling to facilitate Rac1 and RhoA activity and function. Sci. Signal. 2021, 14, eabc0425.
- Tung, J.J.; Kitajewski, J. Chloride intracellular channel 1 functions in endothelial cell growth and migration. J. Angiogenes Res. 2010, 2, 23.
- Lucitti, J.L.; Tarte, N.J.; Faber, J.E. Chloride intracellular channel 4 is required for maturation of the cerebral collateral circulation. Am. J. Physiol. Heart Circ. Physiol. 2015, 309, H1141–H1150.