Your browser does not fully support modern features. Please upgrade for a smoother experience.
Please note this is an old version of this entry, which may differ significantly from the current revision.
Subjects:
Microbiology
The acceleration of climate change has been associated with an alarming increase in the prevalence and geographic range of tick-borne diseases (TBD), many of which have severe and long-lasting effects—particularly when treatment is delayed principally due to inadequate diagnostics and lack of physician suspicion. Moreover, there is a paucity of treatment options for many TBDs that are complicated by diagnostic limitations for correctly identifying the offending pathogens.
- tick-borne diseases
- Lyme disease
- borrelia
- pangenomics
- diagnostics
- borreliaceae
1. Introduction
Even in this era of modern medicine, including mass vaccination achievements and antibiotic treatment regimens, vector-borne diseases such as Lyme borreliosis, Malaria, Dengue fever, yellow fever, and bubonic plague still prevail. This group of ancient and persistent diseases is transmitted to humans through the bite of infected arthropod vectors including mosquitoes, lice, flies, and ticks. These vectors acquire their associated pathogens through blood feedings from multiple hosts throughout complex life cycles. This life-cycle complexity leads to difficulties in diagnoses and treatments, creating a persistent healthcare burden that has resulted in a push to identify biomarkers for the development of improved diagnostics, vaccines, and therapeutics [1].
Research on these fronts has been divided unevenly across different vector species. While a great deal of research has been performed on diseases transmitted by mosquitoes, tick-borne diseases (TBD) are less studied. As their name implies, TBDs are primarily transmitted through the bite of infected ticks. While these small ectoparasitic arachnids have long been documented to cause disease, some as early as 1550 B.C., their true clinical significance was not realized until 1893, when a publication by Smith and Killborne linked Rhipicephalus annaltus to the transmission of the protist parasite Babesia bigemina in cattle [2,3,4,5,6]. Despite this finding, research on ticks and their corresponding TBDs progressed slowly until the latter half of the 20th century.
2. Borreliosis, the Most Common Type of Tick-Borne Disease
The emergence of chronic cutaneous, neurologic, arthritic, and cardiac maladies have been documented worldwide for hundreds of years [31,107,108,109,110,111,112]. In the US, these outbreaks have occurred in waves for centuries. Some of the earliest mentions were documented in Long Island during the early 1600s [113]. These cases were called ‘Montauk’s knees’, ‘Southhampton knee’, or water on the knee due to the arthritic-like symptoms [114]. In the latter half of the twentieth century, another wave occurred at Lyme, Connecticut in the beginning of the 1970s. In this small community, several children presented with numerous non-specific and arthritic-like symptoms. These ailments were later formally recognized as Lyme borreliosis and is commonly referred to as Lyme disease [107,108,109,110,111,112]. Over the years, Lb has continually increased in prevalence within the US and Canada and is now the most common tick-borne disease, accounting for ~500,000 new cases each year in the US alone—due in large part to climate change, deforestation, habitat loss, and loss of predators of the primary mammalian species upon which the ticks feed [115].
In the years that followed the discovery of Lb, its etiological agent, Borreliella burgdorferi, and other Borreliella spirochetes were found to be pathogenic to humans [38,116,117,118,119,120]. Some of these spirochetes do not, however, have the same vectors or pathogenesis. In this instance, other Borreliaceae members can cause Relapsing fever (Rf) and may be transmitted through a tick or louse vector. While these distinctions were made primarily on pathology and geo-locale of origin, later comparative genomic research suggested splitting the genus into two distinct disease-causing genera [121]. The Lb causing spirochetes were then given the new designation of Borreliella, while all other Borreliaceae spirochetes which cause Rf retained its original name of Borrelia.
Lb is a multi-systemic infectious disease with a wide and seemingly unconnected variety of conditions (e.g., polyarthralgias; neurological diseases, including polydysthesias/parathesias, cardiomyopathy, multiple sclerosis, other demylenating diseases, and ataxia; and psychiatric conditions, such as pediatric bipolar disorder and PANS and PANDAS) [122,123,124,125,126,127,128,129,130]. The sole pathognomonic presentation of Lb is erythema migrans, commonly known as the bulls-eye rash. Unfortunately, this presentation may not occur or is not visible to all individuals, occurring in approximately 50% of Lb cases [131,132,133,134]. Lb is divided into distinct stages: localized and disseminated. The disease presentations vary wildly among individuals, as well as by the species of Borreliella. This is illustrated most clearly with common Borreliella spirochetes, B. burgdorferi, B.azfelii, and B. garinii, each of which is endemic to the US or Europe. B. burgdorferi, the most common cause of Lb in the US, is primarily associated with arthritis, while B. afzelii is associated with cutaneous infections and B. garinii with neurological disease in Europe [133].
Due to the spectrum of non-specific symptoms for Lb, diagnoses are often difficult. Currently, clinicians rely on imprecise serological diagnostics and proof of tick-bite before accepting a Lb differential. While the above approaches may be useful in some instances, these current diagnostics have severe limitations, including a highly unreliable negative predictive value. To understand why these diagnostics may fail, it is critical to understand the basic biology of these spirochetes.
Previously, researchers have noted that Borreliellal spirochetes share many features ranging from their obligate parasitic nature within a large network of reservoir hosts, rather organisms that sustain spirochetes and facilitate their reproduction, to dynamic morphology that facilitates their near-constant host invasion [38,135]. Their unique morphology is thought to be created by 11 anti-parallel inter-membrane flagella and a chitobiose peptidoglycan [136,137]. Interestingly, this morphology has been documented to change in response to varying external stimuli [138,139,140,141,142]. It is, however, unclear what the mechanisms underlying the Borreliaceae morphological shifts are.
In addition to altering their morphology as a stress response, Borreliaceace spirochetes can manipulate their host’s immune and inflammatory response to their advantage. This is most clearly seen within I. scapularis ticks where Borreliella spp. reside within the tick midgut. These spirochetes are bound to the tick receptor for OspA (TROSPA) until the initiation of a blood meal [143]. Through feeding, the Borreliella dissociate from TROSPA, then switch their outer membrane surface protein (Osp) composition. The act of feeding induces tick salivary proteins to cover presenting Borreliella OspA and translocate to the tick salivary gland before peritrophic membrane formation [144,145]. Once in the salivary gland, the spirochete can then be transmitted into the new host dermis. During transmission, the Borreliella OspA in the outer membrane decreases and OspC rises [144]. For humans, Borreliella/Borrelia can be transmitted at varying rates depending on the tick species, tick feeding status, microbial strain, and microbial load, e.g., an I. scapularis tick can transmit B. burgdorferi within 24–48 h of initiating blood feeding [146,147,148].
Once Borrelieceae spirochetes have successfully entered the human body, they can persist within the dermis before disseminating. There are two proposed dissemination methods for Borreliella spirochetes: the hematogenous and non-hematogenous routes including the lymphatics or tissue [149]. In both dissemination routes, the spirochetes mitigate the host immune response to prevent recognition by the innate immune system and ultimately delay and distort the development of a T-cell-dependent B-cell response [49,150,151,152,153]. In addition, Borreliaceae spirochetes, can also evade the host immune system through various other means [154]. Both Borrelieceae can utilize the antigenic variation system, vls, present within the genome to evade the complement cascade. Additionally, Borreliella can achieve complement evasion by binding to Factor H, a negative regulator of host complement, to outer membrane proteins CspA, CspZ, and OspE [155,156,157]. Borreliella can also inhibit the classical complement pathway by binding C1r to outer membrane protein BBK32 [158,159]. Through antigenic switching of Borreliella outer membrane proteins, including hypervariable OspC and BBA70, the overall outer membrane composition and pathogenesis of the spirochete can be altered in situ [160,161]. Through these virulence mechanisms, it is believed that if Borrelieceae spirochetes are not successfully cleared by the immune system, they may colonize host tissues to form a persistent infection.
While Lyme borreliosis has become highly prevalent, the impact of elapsing fever (Rf) still remains a significant health concern. The RF-causing Borrelia genus can be transmitted either by ticks or lice around the world [154,162,163,164]. Within the United States, this infectious disease has remained endemic solely within western mountainous regions [164]. Regardless of geo-locale of origin, all variants of Rf have the same symptomology. The illness does have nonspecific symptoms like fatigue, headache, nausea, and muscle/joint aches [165,166]. Important diagnostically, however, is that uniquely induces periodic fever spikes associated with Borrelial septicemias. The fever dissipates during periods of time when there are decreased levels of Borrelia present within the blood but return on a cyclical basis over the course of weeks. Due to the elevated numbers of Borrelia within the blood, Rf is commonly diagnosed through microscopic examination of blood smears (Figure 3).
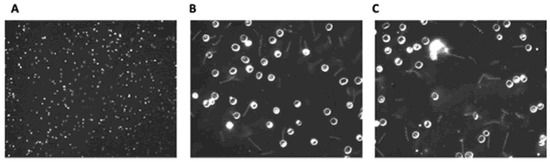
Figure 3. Dark field microscopic images of wet mount Borrelial spirochaetes (Borrelia hermsii strain DAH). Blood from C3H-Hej mice infected with B. hermsii was diluted 4-fold with phosphate buffered saline. (A): 100×, (B,C): 400×.
3. Borreliaceae Diagnostics
In the US, the cost of preventing and treating Lb has been estimated to range from $712 million to $1.3 billion per year, but this is likely a gross underestimate as many Lb patients go undiagnosed for years while seeking care for their ‘nonspecific’ symptoms [167]. Often, patients may pay out of pocket for additional diagnostic tests and treatments. In contrast, acute diagnostics for other common bacteria such as Streptococcus, Treponema pallidum, and Staphylococcus infections are accurate and lead to effective treatment before the bacteria can progress to later stages of infection [168,169,170,171,172]. Meanwhile, the economic impact of Lb infections continues to rise in large part due to an inadequate diagnosis. Thus, it is critical to develop and implement better diagnostics, prognostics, and therapeutics for borrelioses [7,173].
Many factors contribute to acute Lb diagnoses being missed. For example, persons with darker skin pigmentation will often not display a visible EM rash, others lack access to medical care or only have non-specific symptoms of acute Lb, while still others have a non-traditional EM rash that is not recognized during the acute phase of Lb [132,174,175]. If a patient suspected of acute Lb presents to healthcare providers, clinicians will assess risk factors for contracting Lyme borreliosis, including symptom presentation timing, geographic location, recent travel history, pet ownership, and history of other TBDs or rashes [176]. Laboratory-based tests are then utilized by clinicians to confirm a suspected acute case of Lyme borreliosis. Currently, there are several indirect and direct approaches to assist in diagnosing an individual with Lb. The Center for Disease Control and Prevention (CDC) recommends a two-tiered serological (ELISA and Western blot) system to confirm a suspected Lyme borreliosis case [177]. The two-tiered approach relies on a patient’s adaptive immune response towards transiently expressed surface proteins of Borreliella. Producing an IgG antibody response with strong avidity towards specific antigen targets takes between 2–3 weeks following infection [177]. Furthermore, most Lb western blots utilize B. burgdorferi sensu stricto strain B31 (Bb B31) as the source of the proteins utilized in their assay [178]. The B31 subtype was isolated over 30 years ago and the Bb B31 antigens do not represent other Borreliella antigens produced from other closely related Lb causing spirochetes [7,179]. Thus, patients who seroconvert during acute Lb infection could produce antibodies targeting antigens that are not included on the standard western blot. In addition, these serology-based diagnostic approaches cannot serve as prognostics to track treatment outcomes. This forces physicians to primarily rely on a patient’s symptoms to guide clinical outcomes or antibiotic treatment efficacy studies [180].
Serologic Lb diagnostics are further complicated by variation in the human adaptive immune response. If patients are diagnosed with Lb based on an EM rash and antibiotic treatment is promptly initiated, they might not seroconvert [181,182]. This fact further complicates the surveillance and confirmation of Lb. The lack of seroconversion could be due to Bb’s profound immunomodulatory and immunosuppressive effects which depend on the combination of host and pathogen genetics [152,183]. Accurate diagnosis of Lb is further complicated when a patient is co-infected with other Borreliellal spp. or additional tick-borne disease pathogens that are also commonly transferred from the tick’s mid-gut [7,184,185,186,187,188]. Taken together, the average sensitivity of the Lb two-tiered test for the acute Lb is less than 50% [178]. This poor sensitivity produces high rates of false-negatives and delays treatment which contributes to the development of chronic/late-stage Lb. During late-stage Lb, such as Lyme carditis or Lyme arthritis, a two-tiered test can confirm the diagnosis of the patient with high sensitivity. Unfortunately, patients in the later stages of Lb face permanent tissue damage and require longer antibiotic treatments [189,190].
Newer serologically-based diagnostics present recombinantly expressed surface proteins from multiple pathogens and strains of Borreliella [185,191]. These approaches increase the chance of detecting antibodies produced towards B. burgdorferi strains other than B31 or identifying co-infections. These methods, however, still have their limitations, as Bb is immunosuppressive and, depending on the infecting strain and host genetics, a significant percentage of infected persons will fail to appropriately produce antibodies.
Clinicians seeking diagnosis for suspected Lb patients may venture beyond CDC guidelines. Traditional pathologic assays such as dark-field microscopy and primary culture from blood or skin biopsies have poor sensitivity and are not employed as a reliable diagnostic for Lb, however they have a very high positive predictive value [192]. Attempting to culture or detect Borreliella spirochetes from human tissue biopsies using PCR methods (standard or quantitative) is invasive and insensitive [7,193,194,195,196,197]. These direct methods are limited by the low spirochete load in tissues and the bloodstream, unlike many other human bacterial pathologies [7,185,186,198,199,200]. Thus, patients and clinicians require alternative methods of acute Lb diagnosis.
In contrast to the limitations of above-mentioned PCR-based methods for detecting Borreliella DNA within humans, Next-Generation Sequencing (NGS) approaches can be highly specific. As Borreliella spirochetes are rarely present in the blood after initial disease onset, the challenge for NGS is to obtain enough of a sample to confidently detect genes associated with acute Lb bacteria [8,201]. Previously, many Borreliella NGS approaches targeted highly conserved genes throughout the genera such as the ribosomal 16S in ticks or human samples [202]. However, this approach was limited to identifying Lb within ticks rather than humans due to low titers of Borreliella in the bloodstream.
In a new NGS approach, the limitations of the sample sources may be circumvented by using patient urine [203]. While most NGS-based assays are limited by the low counts of Borreliella genomic material present in human samples such as blood, this approach aims to ensure a higher Borreliella DNA yield with claims that Lb bacteria infect the kidneys. While accuracy was stated to be ‘superior’ to the standard two-tiered testing approach, the sensitivity of the test has yet to be reported in the literature. This is slightly different than previous diagnostic iterations which used the same biosample but targeted solely OspA, a protein which would not be expressed in high quantities on the outersurface of Borreliaceae within a mammalian host [204].
Other efforts have been made to increase the sensitivity of Lb bloodborne detection. Traditional PCR-based diagnostics for Lyme borreliosis have been improved by isothermal amplification of DNA, followed by PCR amplification of Borreliellal DNA, which is then detected by electrospray ionization mass spectrometry (PCR/ESI-MSI). In PCR/ESI-MSI, it was possible to detect the presence of B. burgdorferi in 13 of 21 blood samples from patients with an acute Lb cases confirmed with positive serology and a history of at least one EM rash [8,200]. The assay required 1.25 mL of EDTA-treated whole blood and could detect 0.6 or greater copies of Borreliella genomes in whole blood.
In a follow-up study, the PCR/ESI-MSI method attempted to survey the presence of B. burgdorferi sensu stricto within four patients during their antibiotic treatment for Lyme borreliosis [205]. In this study, the investigators increased the blood volume from 1.25 mL to 20 mL, with the aim of increasing the diagnostic sensitivity. B. burgdorferi genes were detected in 2 of the 4 patients acutely infected with the aforementioned spirochete. In addition, they did not determine if the increased blood volume increased sensitivity. The genomic amplification approach relied on detecting and targeting conserved genes present within the Borreliella genome such as rpoC, FlaB, and OspC [206]. While such targets can indicate the presence of this spirochetal genus and may provide species-level resolution, there are some complications [205]. One such complication is that use of consistently expressed proteins like OspC may be insufficient due to the protein’s high diversity.
Another NGS diagnostic approach utilizes unbiased metagenomic cell-free DNA sequencing of human plasma. This cell-free DNA (cfDNA) approach was used to detect B. burgdorferi DNA from 64% (18 of 28) human plasma samples during acute Lb [207]. The cfDNA sequencing method’s sensitivity was further improved by combining the results of the modified two-tiered serology testing to identify 86% of acute Lyme borreliosis cases. Additionally, a recent NGS detection study identified core genes within the Borreliella pangenome to increase the sensitivity of DNA-based Borreliaceae diagnostics [208]. Taken together, genomic Borreliella detection methods have significantly improved over the last decade. However, more work is required to deliver a robust and sensitive diagnosis for patients and clinicians. One possibility is to use innate immune proteins that recognize specific PAMPs, such as Apolipoprotein H linked to paramagnetic beads, to ‘sweep’ a much larger volume of blood [209,210,211,212,213].
Next-generation sequencing has been used to detect host responses to acute Lb rather than attempting to directly detect the Borreliellal genome. Sequencing human T-cell receptors (TCRs) is a novel approach to Lyme disease diagnostics and began clinical trials in 2021 [214]. T-cells respond to Lb infection earlier than B-cells can produce antibodies, and thus the expansion of Borreliellal-specific T-cell receptor sequences in a patient’s circulating lymphocytes has the potential to confirm acute cases of Lyme disease earlier than traditional serologic methods [151]. This TCR immuno-sequencing assay differentiated acute Lb patients from healthy controls with a sensitivity of 54%, while the standard two-tiered serological testing approach had a sensitivity of only 30%. Clearly, human T-cell responses significantly vary between patients and this approach will not detect all acute Lyme disease cases. However, TCR immuno-sequencing assay’s increased sensitivity is a move in the right direction and has the potential to be combined with other diagnostic approaches to further increase sensitivity.
In addition to genomic detection, researchers have explored xenodiagnosis, metabolomics, and biomarker profiling [215,216,217]. Xenodiagnostics use an uninfected, natural vector for the isolation of the targeted pathogen from the infected host [218]. In the case of Borreliellal spirochetes, ticks facilitate the reacquisition of Borreliella from a variety of hosts during a 24-h feeding cycle [219,220]. This feature was noted in the past with Lyme-infected monkeys and mice but, was not substantiated in humans until 2014 [221,222,223]. Over the last 8 years, a clinical trial of tick-based recapture of Borreliellal pathogens from infected human hosts has been underway [218]. No results from this study have been released at this time. This approach could be further bolstered by applying NGS to characterize the pathogens recaptured after the tick feeds on the patient suspected of contracting Lyme borreliosis to increase diagnostic sensitivity. It is important to note, that while it could prove useful, much more work would be necessary to make it feasible as a diagnostic.
Metabolomic analyses of Lyme borreliosis patients has also made great strides in recent years. These studies have identified altered abundances of circulating metabolites produced by host tissues during Lyme borreliosis. A recent assay was able to discriminate between acute Lyme borreliosis and uninfected controls using their metabolic profiles [224,225]. Diagnostics relying on specific metabolic profiles are limited by the time and cost associated to prepare samples for analysis but offer yet another promising avenue for future Lyme borreliosis diagnostics.
Proteomic studies of serum collected from humans afflicted with Lb by Zhou et al. identified host acute phase protein abundance alterations during acute Lyme borreliosis [217]. The abundance of proteins—APOA4, C9, CRP, CST6, PGLYRP2, and S100A9—were validated using a second sample set of acute Lyme borreliosis samples and discriminated between healthy controls and acute Lb patients with a 78% sensitivity. Developing a multiplexed ELISA to identify acute-phase proteins associated with Lyme borreliosis could yield a high-throughput diagnostic, yet the issue of cross-reactivity with other infection markers must first be addressed. “Mimic diseases” such as rheumatoid arthritis or fibromyalgia often have similar acute phase protein alterations. Thus, the Lb proteomic study should be validated for Lb-specificity by testing against a panel of sera from patients with other mimic diseases [224].
Lastly, a glycoproteomic approach using MALDI-FT-ICR mass spectroscopy has been demonstrated to detect changes in the IgG N-glycan profile during acute Lyme disease with a sensitivity of 75% and specificity of 100%. Moreover, this assay can differentiate between acute Lb cases and patients who have received successful doxycycline treatment [226].
In summary, Borreliella diagnostics are improving, but have a long way to go as each has strengths, as well as limitations [178]. In this era of increasing TBDs, the best path forward may be to combine multiple diagnostics to complement the strengths of each method to construct a testing protocol that is highly sensitive and specific. In doing so, better measures can be taken to initiate early treatments and prevent chronic disease progression.
This entry is adapted from the peer-reviewed paper 10.3390/genes13091604
This entry is offline, you can click here to edit this entry!