4. MYCN as Prognostic Indicator in High-Risk Neuroblastoma
Despite the advancement in medical standard therapy for the treatment of HR-NB, there is no specific therapy for MNA patients [
92,
146,
147], but this aspect of the pathology becomes increasingly relevant in consideration of its prognostic effect. For example, the prognostic impact of MNA is particularly relevant in infants with stage M disease where both event-free survival (EFS) and overall survival (OS) are higher in an MYCN non-amplified tumor, that shows better outcomes in comparison with MNA patients (EFS and OS: 82.5% and 90.8% versus 36.9% and 44.8%) [
3]. In a similar manner, the behavior of an L2 and MS stage tumor is mostly impacted by MYCN status. EFS and OS are lower at any age when MYCN is amplified, while the absence of amplification correlates with better survivability results for the patients [
3]. Interestingly, MNA association with worse prognosis appears more pronounced in the context of other favorable prognostic features and can be considered as an indicator for aggressive intervention [
148], highlighting how MYCN gene status impacts therapy selection and medical decision making.
While MNA is considered a staple in neuroblastoma diagnosis, as mentioned before, MYCN over-expression has a more controversial role; in a similar manner, MYCN expression has the same prognostic behavior. Some studies reported that MYCN expression is valuable for prognostic purpose, but only in a specific pathology context. In particular, a patient cohort showed that the lowest value in MNA cases has a more negative outcome when MYCN and mRNA levels are higher, in comparison to patients with higher MNA levels where mRNA expression cannot be considered as prognostic [
11]. Otherwise N-Myc protein expression shows different prognostic value, being reported as an indicator of poor outcome regardless of MNA status [
31]. These findings together suggest that deep investigation on how and when MNA or MYCN expression are predictors of patients’ outcomes needs to be improved, but their value as key elements of prognosis assessment is undeniable, remaining so in the establishment of HR-NB therapy administration.
5. MYCN Determines High-Risk Neuroblastoma
Despite MYCN amplification being the first discovered genetic mechanism in neuroblastoma, its role in driving the pathology is not fully understood [
149,
150,
151]. In fact, MYCN amplification leads to deep remodeling of the cancer cell, influencing its apoptosis resistance, its undifferentiated status, its metabolic landscape and immune evasion.
Different studies showed that MYCN over-expression is an obstacle to neuronal differentiation. High-risk and MYCN amplified neuroblastoma present a different transcriptional profile, where different pathways and genes related to differentiation are particularly altered [
152]. Indeed, MYCN amplified neuroblastoma cell lines fail to differentiate in response to 13-cis-retinoic acid [
8]. The concomitant inhibition of MYCN and the administration of RA is able to reverse this block [
152]. Moreover, the ectopic expression of MYCN in precursor cells blocks the differentiation in chromaffin cells.
In addition, MYCN controls both proliferation and apoptosis: many studies showed that over-expression disrupts the cell cycle, leading to maintained apoptosis inhibition and induced proliferation [
153]. In fact, blocking MYCN leads to G1 phase cell accumulation and slows down the transition to S phase and PI3K repression (which is known to promote cell growth and proliferation) [
154,
155,
156,
157,
158]. MYCN also positively affects the expression of other key cellular regulator such as E2 factor (E2F) and inhibitor of differentiation 2 (ID2), which are also involved in cell cycle progression [
155,
159,
160]. As an additional mechanism, MYCN amplification is also associated to TERT expression and telomere anomalies [
161,
162].
Interestingly, MYCN can promote apoptosis and/or sensitizes cancer cells to cytotoxic drugs [
163,
164]. MYCN is able to promote the expression of phorbol-12-myristate-13-acetate-induced protein 1 (NOXA), which is a pro-apoptotic regulator. Moreover, E-box elements are present in the promoter of p53, which is the most known onco-suppressor able to stop cell proliferation and induce apoptosis (even if in a significant part of neuroblastoma p53 is found mutated) [
164]. As it is known, the murine double minute 2 (MDM2) is a negative regulator of p53 and is over-expressed in different human malignant tumors [
165]. In particular, MDM2 is able to reduce p53 levels using the mechanism of binding to p53 with consequent ubiquitination and proteosomal degradation [
166,
167]. However, MDM2 also promotes the stability of MYCN, while the latter induces MDM2 transcription [
168,
169,
170]. Thus, MYCN can induce the transcription of p53 and MDM2, regulating the balance between proliferation and apoptosis. Over-expression is thought to alter this precarious equilibrium, inducing MDM2 expression and p53 blocking [
168,
169,
170].
Early studies showed how metabolism is deeply altered in cancer cells. In fact, cancer cells are skewed towards rapid ATP production, which is generally obtained through the “Warburg Effect”, where cells rely on glycolysis and mitochondrial respiration is impaired [
171,
172,
173,
174]. Cancer cells then use the fatty acids and glutamine as a source for biosynthesis and ultimately sustain the cell growth and proliferation. Moreover, this alteration in the mitochondria leads to reactive oxygen species (ROS) production, while the fatty acid oxidation is used to replenish the NADPH pool in order to prevent excessive oxidative stress [
175,
176]. Neuroblastoma and, in particular, MYCN amplified tumors are heavily dependent in glutamine, and blocking MYCN leads to the arrest of glutamine transport [
177,
178]. In addition, MYCN promotes the glycolysis and fatty acid uptake and leads to mitochondria alteration [
178,
179]. MNA tumors also present different metabolic alterations, leading to an increase in iron uptake. For instance, it has recently been shown that MYCN induces massive lipid peroxidation and cysteine depletion. This leads MNA to be sensible to oxidative stress and especially to ferroptosis [
180]. In this context, blocking MYCN in MNA neuroblastoma leads to ROS production (through TRAP1 decrease), which the cancer cell fails to handle, consequently undergoing apoptosis [
181]. In addition, MYCN also blocks autophagy and mitophagy (an autophagy sub-pathway used by the cell to recycle damaged mitochondria), and it has been shown in inducible MYCN cell line (TET21N) that the MYCN blocking restores this pathway (OPTN transcription) [
181]. All these studies show a strong rewiring of the metabolism by MYCN expression and the fine grain regulation of the redox equilibrium.
Different studies showed the role of the phosphatidylinositol 3-kinase (PI3K)/mTOR pathway in neuroblastoma [
156,
182,
183]. In fact, the mTOR pathway is known to stabilize N-Myc, and its blocking affects cell growth. Moreover, retinoic acid has been described as being capable of mTOR inhibition. While the N-Myc protein is stabilized by the mTOR complex, it also regulates the expression of different MTOR genes in a positive loop [
152,
184]. Indeed, MNA cell lines have a higher expression of mTOR genes, and they are more resistant to mTOR inhibitors. Furthermore, it has been shown that mTOR is also negatively associated to the prognosis [
152].
MYCN amplification impact is not limited to the cancer cell itself but also to the tumor microenvironment. In fact, MNA cancers remodel the external environment to sustain their growth and the immune evasion [
185]. A significant portion of MNA tumors present PD-L1 expression and MHC I complex down-regulation, leading to a suppressive micro-environment [
186,
187]. In addition, MNAs are also enriched in M2 macrophages and CD4+ T helper 2 cells [
185]. Macrophages are also responsible for maintaining a hypoxic environment and lead to the transcription of hypoxia inducible factor (HIF 2α), which ultimately leads to vascularization and metastasis spreading. Indeed, HR-NB that are fast growing are high in the immunostaining for HIF2 [
188]. However, neuroblastoma also exploits other strategies such as expressing other immune-suppressive molecules (such as CD276), miRNAs and exosomes release [
189]. Overall, both innate and adaptive immune systems seem to be down-regulated [
185]. This complex landscape is probably at the origin of the fact that immune-therapy has shown modest results [
185,
190,
191].
MYCN amplification also leads to extracellular matrix (ECM) modification. ECM is often altered in HR-NB with anomalous collagen I deposit and often correlates with bad prognosis [
192,
193,
194,
195]. These alterations are also promoted by the hypoxic and inflamed state of the tumor microenvironment [
196,
197,
198]. Collagen I inhibition has shown a promising effect, allowing better chemotherapy delivery [
199]. Moreover, different matrix metalloproteinases (MMPs) are altered in neuroblastoma which are linked to bad prognosis, angiogenesis and metastasis promotion [
200,
201,
202].
HIF2 expression, ECM alteration and VEGF expression in HR-NB also lead to new vascularization. These tumors show more aggressive features such as more immature states and more easily spreading metastasis [
203,
204,
205,
206,
207,
208]. Moreover, there is evidence that PI3K kinases promote VEGF expression via MYCN [
209,
210]. Interestingly, blocking PI3K by SF1126 in neuroblastoma led to reduced MYCN expression, cell death and angiogenesis block, while temporarily increasing the macrophages’ M1 to M2 ratio, showing how all these mechanisms are interconnected [
211,
212,
213,
214].
Furthermore, the neuroblastoma tumor microenvironment presents an enrichment of cancer-associated fibroblasts (CAFs) and mesenchymal stromal cells (MSCs). This enrichment correlates with progression and it is important to sustain the tumor growth, micro-vessels formation and progression, showing correlation with poor outcome [
215,
216,
217]. In fact, CAF produces TGFβ (a cytokine with immunosuppressive property) and CCL2, which recruit TAM to neuroblastoma [
218,
219,
220]. In addition, studies have highlighted that the CAF area extension correlates with MYCN amplification [
221,
222,
223]. MSCs are also involved in inducing an immune suppressive environment, leading to the recruitment of ulterior suppressive cells (T regulatory cells and macrophages).
Other than the direct action of MYCN on cellular process leading to neuroblastoma, in the past few years, many epigenetic mechanisms have been discovered regulating MYCN with a specific role in HR-NB development [
224]. Micro RNAs (miRNAs) are small single-stranded RNA molecules that function as post-transcriptional RNA regulators [
225]. MiRNAs targeting MYCN were found to be particularly important in regulating its expression in neuroblastoma in different ways [
8,
226]. Some miRNAs targeting MYCN, such as the let-7 family, work as inhibitors [
227] and are down-regulated in neuroblastoma, inducing N-Myc protein expression [
228,
229,
230]. These miRNA types are considered the most common, but other miRNAs were found to work with the opposite mechanism. For instance, miRNAs such as the family of miR-17-92 are substantially employed as MYCN up-regulators. Interestingly N-Myc is able to stimulate the expression of miR-17-92 cluster, suggesting the presence of a positive feedback mechanism of regulation between MYCN and the miR-17-92 cluster itself [
230,
231,
232].
Moreover, the natural antisense transcript was found to be able to regulate MYCN expression. In particular, MYCN locus is able to generate an antisense transcript known as MYCNOS (or N-cym) [
233]. This transcript originates from the opposite strand of the locus and regulates MYCN as either regulatory long non-coding RNA (lncRNA) or protein. In fact the lncRNA of MYCNOS regulates MYCN promoter through the recruiting of protein in this site [
234], while the MYCNOS-encoded protein works as an inhibitor of glycogen synthase kinase 3 β (GSK3beta), stabilizing in this way the NMYC protein [
235]. For this reason, high levels of MYCNOS can be found relative to MYCN over-expression and correlate with poor outcome in neuroblastoma [
13,
236]. Furthermore, MYCNOS itself was found to be regulated from some non-coding RNAs such as lncUSMycN [
237] that are able to suppress its expression and indirectly regulate MYCN [
234].
Methylation is another regulatory system used by cells to define gene expression and cellular function. While the correlation between MYCN and its methylation status in neuroblastoma is still unclear, many other genes involved in neuroblastoma transformation were well characterized in both MYCN amplified and non-amplified tumors [
224,
238]. Extensive methylation was found in many onco-suppressor genes with no particular correlation to MYCN status [
239]. In particular, miRNAs were reported to be methylated in neuroblastoma cell lines, highlighting their role in tumor progression and poor prognosis [
240].
6. MYCN as Therapeutic Target
Despite current therapeutic advances and ongoing clinical trials, NB remains a complex medical challenge, especially in the high-risk cases, and the discovery of new therapeutic approaches is needed to improve patient welfare and outcome (
Table 2) [
241].
Table 2. Summary of current therapies and the new therapeutic approach in the treatment of neuroblastoma and high-risk neuroblastoma.
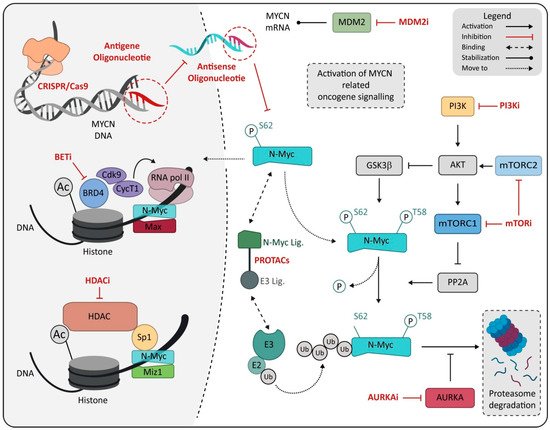
In this scenario, MYCN certainly represents an ideal therapeutic target given its correlation with rapid tumor progression, poor prognosis and the limited expression in normal cells and tissue, suggesting high tolerability for an MYCN-specific approach [
8,
242]. Many attempts and investigations have been made to develop specific inhibitors for N-Myc protein, but both direct or indirect N-Myc modulators (
Figure 2) failed to result in an efficient or reliable N-Myc-specific therapy [
243]. Unfortunately, N-Myc targeting shows a different issue. Some of these issues can resemble any transcription factor, while some challenges are more peculiar and reside in the lack of specific interaction site on the protein [
244,
245] or in the homology with the MYC family oncogene, increasing the difficulty of preserving the physiological function of c-Myc protein in normal tissue [
153,
246]. For this reason, new strategies have been proposed in the hope of overcoming the failure of the precedent attempt to make MYCN a fully available target for HR-NB [
8,
18,
153,
247].
Figure 2. Schematic representation of drugs directly or indirectly targeting MYCN. Drugs are marked in red and reported as class, named on mechanism of action. Abbreviation: MDM2, murine double minute 2; PI3K, Phosphoinositide 3-kinases; PI3Ki, Phosphoinositide 3-kinases inhibitor; BETi, bromodomain and extra-terminal domain family inhibitor; BRD4, bromodomain-containing protein 4; mTORC1, mammalian target of rapamycin complex 1; mTORC2, mammalian target of rapamycin complex 2; mTORi, mammalian target of rapamycin inhibitor; PROTACs, proteolysis-targeting chimeras; MIZ1, MYC-interacting zinc-finger protein 1; HDAC, histone deacetylase; HDACi, histone deacetylase inhibitor; E3, E3 ubiquitin ligase; E2, E2 ubiquitin-conjugating enzyme; AURKA, Aurora kinase A; AURKAi, Aurora kinase A inhibitor; Ub, ubiquitin.