Your browser does not fully support modern features. Please upgrade for a smoother experience.
Please note this is an old version of this entry, which may differ significantly from the current revision.
Endothelial damage is recognized as the initial step that precedes several cardiovascular diseases (CVD), such as atherosclerosis, hypertension, and coronary artery disease. It has been demonstrated that the best treatment for CVD is prevention, and, in the frame of a healthy lifestyle, the consumption of vegetables, rich in bioactive molecules, appears effective at reducing the risk of CVD. In this context, the large amount of agri-food industry waste, considered a global problem due to its environmental and economic impact, represents an unexplored source of bioactive compounds.
- endothelial dysfunction
- agri-food waste
- apple
- pear
- sugar beet
- cardiovascular diseases
1. Introduction
The adult population has an increasing incidence of hypercholesterolemia, diabetes, hypertension, and obesity. Among the major risk factors identified by the World Health Organization, unbalanced eating habits, as well as physical inactivity and high alcohol and tobacco consumption, are the most relevant [1].
Endothelial damage is a crucial aspect of atherosclerosis and precedes the overt manifestation of the disease. In addition, endothelial dysfunction is associated with hypertension, diabetes, ischemia–reperfusion injury, and neurodegenerative diseases [2,3]. It has been demonstrated that the best treatment for cardiovascular disease (CVD) is prevention, and diets containing greater quantities of vegetables appear effective at reducing the risk of CVD [4].
There is a great deal of epidemiological evidence indicating that a diet rich in fruit and vegetables is associated with a reduced risk of many chronic diseases, especially CVD [5,6]. The scientific community is highlighting the important action of natural molecules that can improve biological function and personal well-being [7,8].
Historically, waste from the food chain has been mainly exploited in the production of animal feed [9]. Only in recent decades, however, have these by-products started to be considered a resource to be valorized also in other industrial contexts [10,11]. More recently, studies and projects have begun to analyze and exploit the enormous number of bioactive compounds, still present in by-products, for the development of products with health-promoting properties for human use (foods, cosmetics, supplements, etc.) [12,13]. In this context, by-products from many plants have been studied, demonstrating that they provide health-promoting properties, such as antioxidant and chemopreventive ones [14,15,16].
In northern Italy, traditional apple, pear, and sugar beet crops are still considered among the most high-yielding crops in terms of harvesting, with a large production of by-products during their processing [17].
However, differently from by-products from grapevines, olive, orange, tomato, and chestnut [18,19,20,21,22,23], they are not very well studied for their bioactive content.
2. Apple
According to the Food and Agriculture Organization (FAO), apple production has increased by about 34% in the last 15 years (2006–2020). It was estimated to be about 64 × 106 tons in 2006, while estimation for 2020 was about 86 × 106 tons [70]. China, the USA and Turkey are the largest-producing countries, followed by Iran, India and Italy. In 2020, Italy produced about 2.5 × 106 tons, accounting for about 15.5% of the entire European production [70]. Even though most apple production is destinated to be consumed as it is, about 11.6 × 106 tons/year worldwide is processed to obtain apple products, leading to 3.5 × 106 tons of waste and by-products.
2.1. Exploiting Apple Pomace
Apple pomace is the by-product generated from juice pressing in the production of apple juice and cider. It is a solid mass consisting of a mixture of peel, seed, stem, and pulp residues, and represents up to 30% of the fruit weight. Apple pomace is mainly a source of dietary fiber (50% of dry pomace weight) and phenolic compounds (up to 4 g/kg dry weight) [71]. Apple pomace is still used for animal feeding, and it has very low protein content, which is needed to support animal growth, but several other industrial and agronomical applications have been proposed to exploit these by-products in a circular economy perspective [72]. In the last 25 years, apple pomace has been used to produce several products, such as enzymes, polygalacturonases or hydrolytic depolymerases, involved in the degradation of pectic substances. It has also been applied in food and textile processing [73,74]. Apple pomace has been used as a substrate for bioethanol production [75] and biopolymer synthesis, such as fungal chitosan [72]. More recently, further applications have been proposed for apple pomace. Bioactive compounds from apple waste have been obtained with different extraction methods to be used as food ingredients, in the production of cosmetics, and for their beneficial effects as anti-inflammatory and antioxidant agents in the management of insulin resistance and blood lipid profile [76,77,78,79,80].
Since 2010, chemists have proposed new extraction approaches to improve polyphenol yield from apple pomace, quicken extraction time, and limit the need for organic solvents and purification steps [81]. Attention toward more sustainable and environmentally friendly extraction procedures has pushed the research of new extraction techniques [81,82,83]. Virot et al. in 2010 [82], proposed ultrasound-assisted extraction as a potential method to obtain apple pomace polyphenols. Comparing their procedure to conventional extraction without sonication, they describe their method as able to improve polyphenol yields, more sustainable, less time- and energy-consuming and suitable for upscaling at the industrial level. In terms of yield, ultrasound-assisted extraction led to a +20% catechin extraction, flavan-3-ols and procyanidins increased by 25%, and dihydrochalcones and flavonols by about 5%. In apple pomace, due to both covalent and noncovalent interactions and links with saccharides, polyphenols may result less extractable by traditional methods with aqueous/organic solvents [84]. For this reason, and to improve extraction yields, some authors have applied microwave superheated water technology to apple pomace [85]. With this technique, the polyphenol yield was 5 g/kg dry pomace, with flavan-3-ols, flavonols and dihydrochalcones as the three most present phenol classes [83]. Pulsed electric fields have also been proposed to improve polyphenol yield by increasing cell-membrane permeability through the formation of pores. Advantages of this technique are the reduced extraction time, the possibility to realize bound phenols, leading to a higher total phenol yield, and the low thermal effect [86]. A further extraction approach is represented by enzymatic-assisted extraction. In apple pomace polyphenol-extraction studies, the use of enzymes like endoxylanase and endo-β-1,4-glucanase have been demonstrated to increase phenol yield; however a long time and temperature up to 40 °C are needed [87]. In 2018, Zhang et al. [88] proposed a combination of enzymatic-assisted extraction and ultrasound, defining the optimal extraction conditions to obtain a higher phenol yield. A complete and exhaustive description of polyphenol-extraction methods from apple pomace has been published by Wang et al. [81].
2.2. Cardiovascular Protective Effects of Bioactive Compounds from Apple By-Products
Cardiovascular diseases (CVDs), a wide group of pathologies including atherosclerosis, stroke, cerebrovascular disease, and coronary heart disease, still represent the first cause of death worldwide [89]. Dietary patterns rich in fruit and vegetables providing a significant amount of fiber and phenolic compounds correlate with a lower risk for developing CVDs and CVD mortality. [90]. Clinical trials on apple intake show that apple consumption decrease LDL cholesterol levels [91], improve endothelial function [92], and decrease the body mass index [93].
Phenolic bioactive molecules mediate cardiovascular protection by different mechanisms, such as direct and indirect antioxidant effect [5], anti-inflammatory activity [94], and maintenance of nitric oxide (NO) homeostasis by modulating endothelial nitric oxide synthase (eNOS) activity.
Waldbauer et al. [95] evaluated the ability of polar and apolar fractions of apple pomace methanol/water extracts to modulate eNOS activity in a human endothelium-derived cell line (EA.hy926). They found that the mixture of triterpenoic acids, instead of total phenolic fraction content, was directly related to eNOS activation, suggesting that apple pomace extracts can improve endothelial function. In further research, both on human vein endothelial cell (HUVEC) cultures and in a human clinical trial, the effect of apple polyphenols on vascular oxidative stress and endothelial function was studied [5]. Cell culture studies demonstrated the ability of glycosylated phloridzin, chlorogenic acid and quercetin from apple skin to counteract uric acid production in a dose-dependent manner by inhibiting xanthine oxidase. Moreover, the clinical trial demonstrated that 300 mg/day of apple polyphenols for 8 weeks reduced fasting glycemia, serum uric acid and improved endothelial reactivity [5]. Other studies have demonstrated the possibility to modulate glucose absorption, to reduce postprandial glycaemia and insulinemia by phloridzin, chlorogenic acid and quercetin treatment. Phloridzin and quercetin are known to inhibit, in vivo, the intestinal glucose transporters SGLT1 and SGLT2 [5,96,97].
Apple phenols have been claimed to exert cardiovascular protective effects by acting as antioxidants. In fact, phloridzin and quercetin are known to exert indirect antioxidant effects by inducing gene expression of phase-2 detoxifying and antioxidant enzymes [98,99]. Quercetin is known to protect cardiac and endothelial cells from oxidative stress, and in cardiac cells quercetin was demonstrated to upregulate gene expression of phase 2 enzymes, highlighting its ability to act with an indirect antioxidant mechanism, and to modulate PI3K/Akt and ERK1/2 signaling pathways [100,101]. In a model system represented by human brain, microvascular endothelial cells exposed to hypoxia and reoxygenation, quercetin exerted its antioxidant effect by activating the nuclear erythroid 2-related factor 2 (Nrf2) signaling pathway [102].
Nrf2 induction was also the molecular target involved in apple pomace extracts that induced antioxidant effect in a mouse model of liver oxidative stress [103].
Inflammation and oxidative stress have been defined as “inseparable partners” not only because the chronic or excessive activation of one of the two phenomena inevitably determines the activation of the other but also because the molecular mechanisms and pathways involved cannot be untangled and they share a mutual signaling cascade [104]. Apple polyphenols exhibit anti-inflammatory activity not only in in vitro studies but also in animal models and in clinical trials of chronic inflammatory diseases [105]. In obese subjects, postprandial apple intake reduced plasma level of proinflammatory cytokines IFN-γ, IL-6, and TNF-α [106]. In vitro, quercetin exerted an anti-inflammatory effect by inhibiting LPS-induced iNOS protein expression and subsequent NO overproduction in cardiomyoblasts (H9C2 cell line) [107]. However, administration of quercetin (500 mg/day) to post-myocardial infarction patients did not result in a significant reduction of proinflammatory biomarkers [108], highlighting that administration of a single purified bioactive compound does not always provide positive effects. Eleven pentacyclic triterpenes have been detected in apple pomace [95], and among them, tormentic and ursolic acid have been shown to reduce inflammation by inhibiting NF-κB signaling [109,110].
Apple pomace is not merely a source of phenols—50% of its mass is represented by dietary fiber. Even though fibers cannot be considered bioactive compounds, they play a role in the prevention of CVD-related risk factors [111]. Apple fibers are both insoluble, e.g., cellulose and hemicellulose, and soluble, e.g., pectin, which in turn contains homogalacturonans and rhamnogalacturonans. Galacturonic acid residues are partially esterified, methylated, and the degree of methylation appears to modulate viscosity and increase the amount of cholesterol eliminated through the feces [112,113]. Before being fermented into short-chain fatty acids by gut microbiota, apple pectin modulates bile acid enterohepatic circulation and reduces plasma cholesterol [114,115].
An apple cholesterol-lowering effect has also been investigated in human studies. Ravn-Haren et al. (2013) evaluated the effect of daily apple, apple pomace, or juice consumption for 4 weeks on total and LDL cholesterol levels in a small group of healthy subjects. Even if nonsignificant, after the dietary treatment period they observed a trend of cholesterol reduction [116]. More recently, Koutsos et al. [117] evaluated in 40 human subjects the cholesterol-lowering effect of the consumption of two apples/day for 8 weeks and demonstrated that the treatment was effective in reducing serum, total and LDL cholesterol, and triglycerides.
Apple fibers are not the only apple component responsible for reducing cholesterol levels. They may both bind directly to bile acids, increasing their excretion in feces and activating hepatic cholesterol 7alpha-hydroxylase increasing cholesterol excretion, as was suggested by supplementation studies in an animal model [118]. A representative summary of the cardioprotective role of apple pomace is reported in Figure 2.
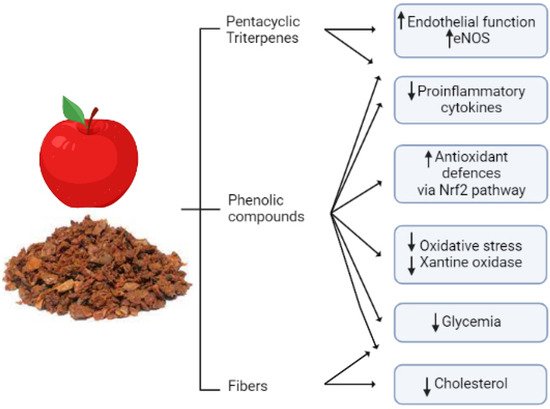
Figure 2. Apple pomace classes of bioactive compounds and their cardio- and vasculoprotective actions. Apple pomace contains fibers that mainly provide cholesterol-lowering effects, but it also contains bioactive compounds such as phenols and triterpenes that provide multiple effects positively affecting the cardiovascular system, such as improvement in endothelial function and antioxidant effect, reduction in proinflammatory cytokines and oxidative stress, and a hypoglycemic effect. Overall, thanks to its composition, apple pomace exerts pleiotropic cardio- and vasculoprotective effects by targeting multiple signaling pathways and risk factors. The figure is original and created with BioRender.com.
3. Pear
Data from the FAO describe that worldwide pear production increased by 15.6% between 2006 and 2020. Most recent available data show that world pear production was about 23 × 106 tons in 2020, with China, Italy, and the USA the largest-producing countries. In the last 15 years, pear production in Italy has decreased by about 32%; however, Italy is still the top producer in Europe and the second worldwide [70]. It is one of the most important stone fruits and widely consumed as fresh when fully mature and/or in processed forms, such as in purees and jam. The other pear parts, such as peels, seeds, and leaves, are usually discarded.
3.1. Exploiting Pear Pomaces
As well as apple pomace, pear pomace is the by-product generated from pear processing and it still contains bioactive compounds, carbohydrates and fibers that can be exploited at the industrial level. Possible applications of pear pomace have been proposed: as a substrate for bacterial cellulose and pear vinegar coproduction [119], as a source of fiber and antioxidants for the prevention of obesity and oxidative stress in animal studies [120,121].
The analyses of phenolic compounds in different anatomical pear parts demonstrated that pulps contain the lowest concentration of phenolic bioactive molecules, which, in turn, are enriched in seeds and peels [122]. For this reason, pear pomace appears a good source of these bioactive compounds. Characterization of pear bioactive molecules has been carried out, demonstrating the presence of polyphenols such as phenolic acids and flavonoids, and triterpenes [123]. In 2014, Li et al. [124] analyzed the chemical composition of peels and pulp from 10 pear varieties. They found that, both in peel and pulp, the prevailing monomeric compounds were arbutin, oleanolic acid, ursolic acid, chlorogenic acid, epicatechin, and rutin. Moreover, quantitative analyses of total phenols, flavonoids, and triterpenes content revealed that all compounds detected were 6–20 times more concentrated in the peel than in pulp [124], suggesting that in the pear-processing food chain most bioactive compounds are discarded in pear pomace.
3.2. Cardiovascular Protective Effects of Bioactive Compounds from Pear By-Products
Pear bioactive molecules have been claimed to exert multiple healthy effects. At the cardiovascular level, they might be beneficial by positively modulating numerous risk factors for cardiovascular diseases. Chlorogenic acids have been demonstrated to exert anti-inflammatory properties by downregulating TNF-α and IL-8 in cell culture studies [125]. Pyrus pashia, known as the Himalayan pear, extracts, rich in chlorogenic acid, catechin, epicatechin, and arbutin, inhibited 5-lipoxygenase, cyclooxygenase-2 activities and reduced IL-6 and TNF-α expression in vitro. Moreover, these extracts demonstrated similar anti-inflammatory properties in an in vivo mouse model [126]. Oleanolic acid, a pentacyclic triterpenoid detected in pear peels, is known to exhibit anti-inflammatory properties. In a model system of carotid artery-injured diabetic rats, oleanolic acid suppressed the inflammasome signaling pathways and reduced serum levels of TNF-α, IL-1β, IL-6 and IL-18 [127]. A recent study on olive oil pomace, which is also rich in oleanolic acid, highlighted the anti-inflammatory properties of this triterpenoid [128].
Studies focused on arbutin biological effects showed its antioxidant and anti-inflammatory properties. In cardiac cells (H9C2), arbutin pretreatment protected cells from isoproterenol-induced oxidative stress [129]. Moreover, in an in vivo model system of cardiac damage induced by lipopolysaccharide in rats, arbutin treatment reduced inflammation by modulating TNF-α and IL-6 levels [130].
Pear peel components, such as chlorogenic acid, have been proposed to exert direct effects on the vascular endothelium. Ex vivo and in vitro studies showed that chlorogenic acid improved NO production at the vascular level and partially inhibited angiotensin converting enzyme (ACE)-I [131,132].
Pear and pear by-product extracts have also shown antihyperglycemic properties. Both in rats and diabetic mice, pear peel extracts reduced fasting glycemia. Different mechanisms have been proposed to explain this effect: on one side, these extracts seem to slow down carbohydrate digestion by inhibiting alpha-amylase and alpha-glucosidase activity; on the other, they have been claimed to stimulate pancreas insulin secretion [133,134].
Pear peal extracts have also been claimed to modulate hyperlipidemia, a further risk factor for CVDs. In a mouse model, this extract lowered triglycerides, total cholesterol, and LDL [134]. A representative summary of the cardioprotective role of pear pomace is reported in Figure 3.
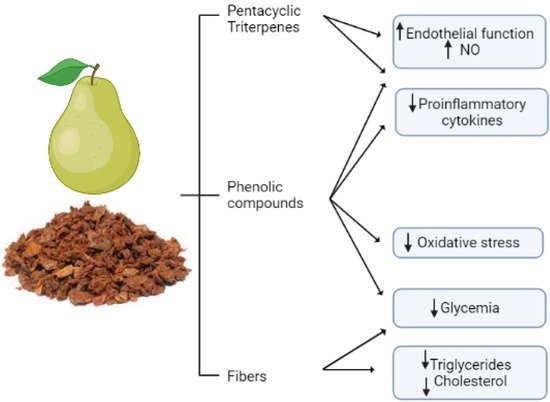
Figure 3. Pear pomace classes of bioactive compounds and their cardio- and vasculoprotective actions. Pear pomace contains fibers that mainly provide cholesterol- and triglyceride-lowering effects, but it also contains bioactive compounds, such as phenols and triterpenes, that provide multiple effects positively affecting the cardiovascular system, such as improvement in endothelial function, reduction in proinflammatory cytokines and oxidative stress, and a hypoglycemic effect. Overall, thanks to its composition, pear pomace exerts pleiotropic cardio- and vasculoprotective effect by targeting multiple signaling pathways and risk factors. The figure is original and created with BioRender.com.
4. Sugar Beet
Sugar beet (Beta Vulgaris variety Saccharifera) is one of the most important crops, along with cane, for sugar production in Europe [135]. Nowadays, 120 million tons of beets are produced in Europe, from which about 16 million tons of sugar are extracted (50% of European production of sugar). Even if sugar beet is one of the traditional crops produced in Italy, only 30% of its national demand is covered, while 70% comes from abroad, mainly from France and Germany, which are the leading producers. The major sugar beet-producing regions in Italy are Emilia-Romagna, Veneto, Puglia, Lombardy, and Marche.
The cultivation of beet is one of the most virtuous in terms of greenhouse gas emissions and it is strategically relevant in agricultural rotation thanks to its ability to release organic nutrients and minerals into the soil. It is considered an “improving” crop capable of enriching fields after harvest, increasing their production yield, and “renewal” for tillage and fertilization at the beginning of the rotational cycle. When introduced into the rotation process, it significantly reduces the environmental impact of farming [136,137]. As such, the sugar beet plant has not just the peculiar characteristic of accumulating sucrose in the root system but can be seen as a fundamental crop for agricultural sustainability. Therefore, this plant is of particular interest not just from an economic point of view.
4.1. Sugar Beet Waste and By-Products
The primary aim of growing sugar beet is the production of refined sugar. However, any other by-products that remain or are produced in the manufacture of refined sugar from beets, i.e., beet tops (leaves and crowns), pulp, and waste molasses, might have a much greater commercial value than the original one. Right now, the alcohol made from waste molasses and commercial fertilizers made from refuse slop represents an effective “upcycling” way to convert a by-product into a high-added-value one in economic terms. Attempts are also being made to use beet leaves in the production of methanol, so general interest may be taken in the proper utilization of the sugar beet by-products. Here, we discuss the main bioactive molecules present in sugar beet products and their possible beneficial role in endothelial function (Figure 4).
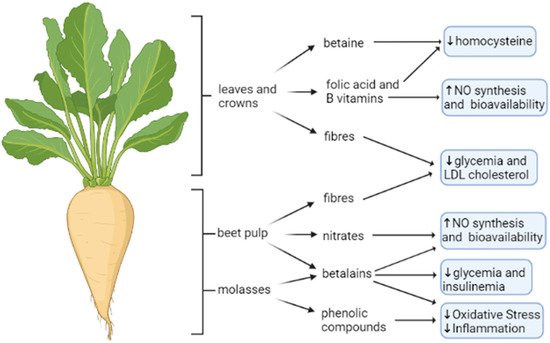
Figure 4. Sugar beet bioactive compounds and their cardio- and vasculoprotective actions. The entire sugar beet plant is a source of bioactive molecules with a potential cardio- and vasculoprotective role. Betaine and B vitamins from leaves and crown are involved respectively in homocysteine detoxification and NO synthesis. Nitrates, betalains and phenolic compounds from beet pulp, and its by-product molasses, are involved in NO bioavailability, glycemia control and reduction in oxidative stress and inflammation. Moreover, fibers from different plant parts exert hypoglycemic and cholesterol-lowering effects. The figure is original and created with BioRender.com.
The first by-product obtained in sugar beet processing is the top, made of leaves and crowns, which is removed by the grower in preparing the beets for the factory at harvest time. Although the sugar is produced in the leaves, it constantly passes into the root, where it is stored. Sugar beet leaves also contain betaine, fiber, folic acid, and B vitamins [138].
Beet leaves have a huge quantity of the amino acid betaine, used by plants as an organic osmolyte to protect against osmotic stress, aridity, high salinity, or temperature changes [139]. The intracellular accumulation of betaine, by not disturbing enzyme functions, protein structure, or membrane integrity, allows water retention in cells, protecting them from the effects of dehydration. Betaine is also a methyl-group donor, used by cells as a cofactor in the biochemical process called “transmethylation” which is essential for cellular metabolism. Betaine plays a key physiological role in the process of detoxifying homocysteine (Hcy), a powerful oxidant, into methionine. Recently, a QuEChERS (quick, easy, cheap, rugged and safe) procedure was successfully applied for the isolation and quantification of betaine in different beetroot parts (peel, juice, pulp), revealing betaine levels from 2 to 5 mg/g according to the plant fraction [140].
The sheer volume of extracted waste from sugar production is molasses and pulp. Indeed, just the fresh pulp constitutes about 80% of the weight of the beets, and following the extraction of sucrose, the pulp is separated off and frequently used worldwide in animal feed or converted into biogas, while molasses are recycled for livestock feed and alcohol-based preparations [141,142]. Fibers in sugar beet root crops are not extensively lignified; generally, they comprise approximately one-third pectin, one-third hemicellulose, and one-third cellulose, so these by-products can have functional characteristics due to their significant fiber content, both soluble and insoluble, which can provide interesting technological properties for the food industry, such as enrichment of pasta, cakes, and cookies [143,144].
High amounts of antioxidant molecules are found in sugar beet molasses [145], which means that this by-product could be used as a raw material for the production of functional foods, nutraceuticals, and supplements. Molasses is the by-product that remains after the crystallizable sugar has been separated from the concentrated beet juice, so it contains nearly 50% of sugar that cannot be separated from the non-sugars by ordinary methods, owing to the presence of various salts that have been taken up by the beet from the soil in the process of growth [146]. Chen and coworkers [145], recently observed through HPLC-DAD-MS/MS a high quantity of anthocyanins (cyanidin-3-O-rutinoside, cyanidin-3-O-glucoside, delphinidin-3-O-rutinoside, and delphinidin-3-O-glucuronide) in sugar beet molasses extracted by an eco-sustainable and chemical-free ultrasound-assisted process (UAE). They also detected other functional components with known antioxidant properties, such as gallic acid, vanillin, hydroxybenzoic acid, syringic acid, catechin, and ferulic acid.
This entry is adapted from the peer-reviewed paper 10.3390/antiox11091786
This entry is offline, you can click here to edit this entry!