Your browser does not fully support modern features. Please upgrade for a smoother experience.
Please note this is an old version of this entry, which may differ significantly from the current revision.
Subjects:
Biochemistry & Molecular Biology
Increasing environmental distress is associated with a growing asthma incidence; no treatments are available but montelukast (MTK)—an antagonist of the cysteinyl leukotrienes receptor 1—is widely used in the management of symptoms among adults and children. Initially described as the slow-reacting substances of anaphylaxis, leukotrienes (LTs) are pro-inflammatory lipid mediators derived from arachidonic acid.
- montelukast
- leukotrienes
- adverse drug reactions
1. Introduction
Montelukast (MTK) is an antagonist of the cysteinyl leukotrienes receptor 1 and is routinely used in the management of asthma symptoms among adults and children. Its systemic anti-inflammatory actions, which are particularly important in the brain tissues, are at the onset of various clinical studies focused on the repurposing of this drug for various other diseases, aimed particularly at Alzheimer’s and Parkinson’s diseases. However, this repurposing clashes with neuropsychiatric adverse drug reactions elicited by the drug.
2. Cysteinyl Leukotrienes—Multifunctional Inflammation Mediators
2.1. Cysteinyl Leukotrienes and Their Receptors
Initially described as the slow-reacting substances of anaphylaxis, leukotrienes (LTs) are pro-inflammatory lipid mediators derived from arachidonic acid [1,2]. These mediators are synthesized mainly in cells from the innate immune system (e.g., polymorphonuclear leukocytes, macrophages, mast cells, and brain microglia) following activation by immune and non-immune stimuli such as infection, tissue injury, allergens, and exercise. Upon cell activation, the cytosolic calcium concentration increases, and the cytosolic phospholipase A2 (cPLA2) and 5-lipoxygenase (5-LOX) enzymes are activated and translocated to the nuclear envelope. There, cPLA2 cleaves glycerophospholipids, releasing arachidonic acid (AA), which is converted to the acyclic hydroperoxide 5(S)-hydroperoxyeicosatetraenoic acid (5-HpETE) by 5-LOX-mediated oxidation upon LOX activation by 5-LOX activating protein (FLAP); 5-HpETE, in turn, undergoes dehydration to the unstable conjugated triene epoxide leukotriene A4 (LTA4), the first metabolite in the leukotriene pathway. LTA4 is a short-lived intermediate that can undergo conjugate addition of water to form leukotriene B4 (LTB4) or conjugation with glutathione by LTC4 synthase to form leukotriene C4 (LTC4, an S-glutathionyl LT). LTB4 and LTC4 are transported to the extracellular space mainly by multidrug resistance proteins, namely through MRP4 (LTB4) and MRP1 (LTC4) [3,4], where cleavage of LTC4 to leukotriene D4 (LTD4) and subsequently to leukotriene E4 (LTE4) takes place. LTD4, an S-cysteinyl LT, is synthesised from LTC4 by a γ-glutamyl transpeptidase (GGT)-mediated cleavage, whereas LTE4 results from the cleavage of LTD4 by a membrane-bound dipeptidase [5,6,7,8,9,10,11,12,13,14,15,16,17,18].
LTB4 is a pro-inflammatory LT that acts on human polymorphonuclear leukocytes (PMNLs) such as neutrophils, via G protein-coupled receptors B-LT1 or B-LT2, triggering chemotaxis and the subsequent activation of the inflammatory response. LTC4, LTD4, and LTE4 constitute a group of cysteinyl leukotrienes (CysLTs) that act through G protein-coupled cell surface receptors, of which the two classical receptors are the cysteinyl leukotriene receptors 1 (CysLTR1) and 2 (CysLTR2). LTC4 is an agonist of CysLTR1 whereas LTD4 binds CysLTR1 and CysLTR2. LTE4 is described as an agonist of CysLTR3 (also known as GPR99 receptor) and of the purinergic receptors GPR17 and P2Y12 [5,6,7,8,9,10,11,12,13,14,15,16,17,18].
Cysteinyl leukotriene receptors (CysLTRs) are involved in the pathophysiology of various respiratory allergic diseases, including bronchial asthma, exercise- and aspirin-induced asthma, and allergic rhinitis, as well as atopic dermatitis, allergic conjunctivitis, and anaphylaxis, exhibiting a large overlap with the B-LT receptors, but allowing a finely tuned immune response [11,12,13,20,21]. Receptor engagement by CysLTs promotes bronchoconstriction, vascular leakage, and neutrophil extravasation to inflammation sites [7]. CysLTR1 is expressed in most human tissues, particularly in the appendix, oesophagus, gall bladder, lung, lymph nodes, spleen, and urinary bladder. The affinity of leukotrienes to this receptor varies in the order LTD4 > LTC4 > LTE4. This receptor is sensitive to classical antagonists (Figure 2) such as montelukast (MTK, Singulair®), zafirlukast (Accolate®), pranlukast (Onon®, Azlaire®), pobilukast, and MK571, all members of the Lukast group (cysteinyl leukotriene receptor antagonists).
CysLTR2 is predominantly expressed in the spleen, heart, brain, and adrenal gland, and its affinity strength is LTC4 = LTD4 > LTE4. HAMI3379 (Figure 2) was identified as a potent and selective CysLTR2 receptor antagonist [22]. To our knowledge, only two dual inhibitors of both CysTR1 and CysLTR2 are reported—BAY-u9773 and gemilukast (Figure 2). However, BAY-u9773 is neither very potent nor selective for human CysLTs [11,12,20,21,23] and gemilukast did not show outcome differences when compared with MTK [24,25]. Figure 2 also shows the experimental IC50 values available for these compounds.
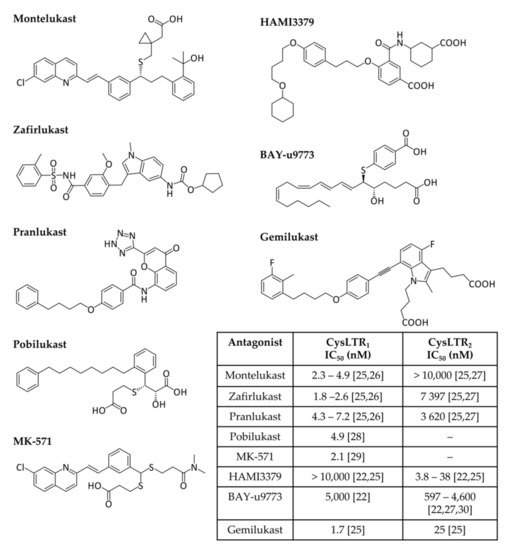
Figure 2. Cysteinyl leukotrienes receptor antagonists. CysLTR1 antagonists (montelukast, zafirlukast, pranlukast, pobilukast, and MK571), CysLTR2 antagonists (HAMI3379) and dual antagonists (BAY-u9773 and gemilukast) are shown. Experimental IC50 values available from the literature are also given in the inset table [22,25,26,27,28,29,30].
Besides these classical receptors, three other receptors are associated with the leukotriene cascade—GPR99, P2Y12, and GPR17.
GPR99, or OXGR1, is an α-ketoglutarate receptor that was originally thought to be a P2Y receptor [31]. This receptor is expressed in the kidney, placenta, trachea, salivary glands, lungs, and smooth muscle cells, as well as in some brain regions; in addition to its effects on acid–base homeostasis, it is also involved in axon growth [32,33,34,35]. GPR99 is considered the third CysLT receptor (CysLTR3) due to its high affinity for LTE4. No antagonists are currently available for this receptor [32,36].
P2Y12 is an adenosine diphosphate receptor that also mediates LTE4-dependent pulmonary inflammation (but not the LTD4 response) [37]. This receptor is mainly expressed in platelets and microglia, where it triggers platelet activation and blood clotting, and induces microglial chemotaxis in situations of central nervous system (CNS) injury [38,39,40,41]. P2Y12 is also associated with some asthma symptoms, namely with eosinophilic inflammation and airway hyper-responsiveness [42,43]. The P2Y12 receptor is blocked by anti-platelet drugs such as clopidogrel, prasugrel, and ticagrelor [44].
Lastly, GPR17 is a uracil nucleotide P2Y receptor expressed in the brain that also binds CysLTs [12,14,45,46,47,48,49,50]. This receptor is described as a sensor of neuronal damage, being activated by nucleotides and CysLTs released in the damaged area and plays a dual role depending on its surroundings: under physiological conditions, GPR17 contributes to the differentiation and maturation of oligodendrocytes, whereas under pathological conditions it mediates demyelination and apoptosis [51,52,53,54,55,56]. GPR17 is described as a putative negative regulator of CysLTR1 [57]. The CysLTR1 inhibitors pranlukast and montelukast are also antagonists of this receptor [46,58,59].
2.2. Leukotrienes in the Brain
The potential of leukotrienes as pro-inflammatory lipid mediators, described above, together with the pattern of expression of their receptors in different organs, has led to the suggestion that LTs play an important role in the central nervous system. In fact, recent advances have associated inflammation with some brain pathologies such as multiple sclerosis, Alzheimer’s disease, Parkinson’s disease, brain ischemia, and epilepsy, among others, and leukotrienes are thought to play a role in this process [60,61].
Despite having been originally found in leukocytes, leukotrienes are also present in the brain. Not only is the 5-LOX enzyme widely distributed in various brain regions (e.g., cortex, hippocampus, and cerebellum), but CysLTs are also produced by vascular endothelial cells, neurons, and glial cells upon LTA4 expression by activated neutrophils [47]. CysLTR1 is widely expressed in the cortex, hippocampus, and nigrostriatum, as well as in cerebrovascular endothelial cells, astrocytes, microglia, and several types of neurons. On the other hand, CysLTR2 is expressed in the cortex, hippocampus, substantia nigra, astrocytes, microglia, and neurons [62,63,64,65,66]. These receptors are usually weakly expressed unless activated by pathological stimuli [66]. Some studies have shown that the exposure of neurons to acute neuronal injury is associated with upregulated levels of CysLTR1 and CysLTR2, and with increased blood–brain barrier (BBB) permeability. Once activated, CysLT receptors will trigger an inflammatory cascade, activating pro-inflammatory cytokines and inflammation, ultimately leading to neuronal damage [62,64,65,67].
2.2.1. Leukotrienes: Role in Neuroinflammation
Neuroinflammation is a complex biological response of the brain and spinal cord mediated by the production of pro-inflammatory cytokines (IL-1β, IL-6, and TNF-α), chemokines (CCL2, CCL5, and CXCL1), reactive oxygen species (ROS), and other mediators (NO, prostaglandins, and leukotrienes) [68,69,70]. This biological response is associated with restoration of homeostatic balance, in order to eliminate and repair the initial cause of cell injury, and can be classified as acute (seconds to days) or chronic [68,71].
An acute inflammatory response is an adaptive response, usually beneficial, meant to protect tissues from a specific injury as trauma or infection [71]. In situations of acute inflammation, the immune system priorities are neuroprotection, tissue repair, and neuroplasticity. When the brain is exposed to immune signals after any infection, microglia and astrocytes are activated and neuroinflammatory cytokines such as IL-1β, TNF-α, and IL-6 are expressed to sustain the inflammatory response. This response is short and transient, and no severe effects take place [68]. Brain development and plasticity are other positive aspects of neuroinflammation. Neurons, astrocytes, and glia cells are involved in neurotransmission through the modulatory effect of cytokines and neuromodulators such as IL-1β, IL-6, TNF-α, NF-κB, and glutamate [68,72]. Brain tissue repair can also be stimulated through the activation of macrophages, lymphocytes, and microglia, which promotes angiogenesis, axon regeneration, myelin clearance, and oligodendrocyte regeneration [68,73,74,75]. Lastly, immune system training through immune pre-conditioning or euflammation allows modulation of the microglia response against hyper-inflammatory conditions, protecting the brain from CNS injuries [68,76,77].
However, if the acute inflammation response fails and the inflammation process persists, chronic inflammation ensues with a long-lasting maladaptive or defective response that could destroy tissues and compromise the immune response [70,71]. Characteristically, there is an increased production of cytokines (IL-1 and TNF-α), reactive oxygen species (ROS), and other inflammatory mediators (e.g., inducible nitric oxide synthase, iNOS), associated with the activation of microglia cells, and consequent expression of more pro-inflammatory cytokines and chemokines in the brain [68]. This activation could be caused by noradrenergic signalling, inflammasome activation, and ATP release [78,79,80]. Microglia activation is also involved in the recruitment of monocytes from the bone marrow to the brain and is linked to anxiety-like behaviour and to the development of mood disorders [68,81].
The normal ageing process is one example of the disruption of the communication pathways between the brain and the immune system, leading to chronic neuroinflammation. During ageing, there is an increase in inflammatory (e.g., IL-1β and IL-6) and a decrease in anti-inflammatory (e.g., IL-10 and IL-4) cytokines that results in damage to the nervous system and the onset of neurodegenerative diseases [68].
It has been shown that the leukotriene receptors CysLTR1 and CysLTR2 in different brain cells, namely microglia (known as the brain’s immune system), astrocytes, and several types of neurons, are upregulated in response to brain injury such as brain ischemia, Alzheimer’s disease, and Parkinson’s disease [62,63,64,65,82,83,84,85,86,87,88]. The modulation of these receptors is associated not only with the outcome of acute inflammation but also with the restoring of homeostasis during chronic inflammation [62,63,64,65,82,83,84,85,86,87,88].
Although the mechanisms of action are still poorly understood, evidence supports the relationship between leukotrienes and neuroinflammation, suggesting the use of leukotriene antagonists as a possible therapeutic strategy in neuroinflammation, given that antagonists of either CysLTR1 or CysLTR2 display wide multi-target anti-inflammatory activity [66]. Both receptors are expressed at low levels in multiple brain regions, but are upregulated following injury, as observed in various experimental models of ischemia and Alzheimer’s and Parkinson’s diseases [65,82,83,84,89]. Interestingly, silencing the expression of the genes coding for these two receptors leads to in vivo protection against lipopolysaccharide- and ischemia-induced brain inflammation and injury [87,88]. Although this strategy needs to be further explored, it could be a very promising therapeutic approach to the improvement of symptoms (or even disease treatment) in patients who suffer from neurodegenerative disorders and have no alternative therapy to manage the debilitating symptoms characteristic of neurodegeneration.
2.2.2. Leukotrienes in Neuro-Signalling Pathways
Message transmission between neurons results from an electrical impulse (action potential) that causes the release of neurotransmitters into the synaptic cleft. After crossing the synaptic cleft, neurotransmitters will reach their receptors on the postsynaptic side to excite or inhibit the target neuron. Excitatory synaptic transmission is mainly assured by L-glutamate, whereas γ-aminobutyric acid (GABA) is the major neurotransmitter involved in the inhibitory synaptic response. In addition to these neurotransmitters, there are other molecules involved in signalling and neuromodulation, such as acetylcholine, monoamines (e.g., dopamine, adrenaline, serotonin, and histamine), purines (e.g., adenosine), and neuropeptides [89].
A close relationship between neuroinflammation and neuro-signalling pathways has been proposed. One example is the involvement of excitotoxicity in neuroinflammation: an exacerbated or prolonged activation of glutamate receptors, particularly the N-methyl-D-aspartic acid receptors (NMDA), causes an increase in calcium influx into the neurons. This increase of intracellular calcium levels leads to a neurotoxic response, including the activation of the AA pathway, that can lead to the loss of neuronal function and, ultimately, cell death [90]. Studies involving CysLTR antagonists showed that pranlukast was able to inhibit NMDA-induced CysLTR1 expression, leading to a decrease in excitotoxic cell death [91]. Montelukast also presented a strong anti-excitotoxicity effect, as well as anti-inflammatory and neuroprotective properties [83].
Dopamine reuptake is also associated with the leukotriene pathway. Inhibition of the 5-LOX activating protein (FLAP) is associated with the improved integrity of dopaminergic neurons [92].
2.2.3. The Leukotriene Link between Stress and Depression
Depression can result from chronic neuroinflammation. Not only pro-inflammatory cytokines (e.g., IL-1β and TNF-α) were found to be dysregulated in depression patients, but also IL-1β, IL-6, TNF-α, or lipopolysaccharide (LPS) administration in animal models led to depression- and anxiety-like behaviours [93,94,95,96].
Stress stimuli led to an increase in calcium concentration, releasing AA after cPLA2 activation by phosphorylation [97]. Once released, AA is used to synthesise leukotrienes and prostaglandins. A study using mice in which the cysltr1 gene was silenced in the hippocampus suggested that the absence of CysLTR1 prevents the development of neuroinflammation and of a depressive-like phenotype [98]. The effects observed upon blocking the same receptors in a mouse lipopolysaccharide-induced neuroinflammation model support those previous results [99]. Inhibition of the 5-LOX enzyme has also been associated with a relief of depression-like behaviour [100].
2.2.4. The Role of Leukotrienes in Neurodegenerative Diseases
Besides their role in inflammation, leukotrienes are also involved in some of the most characteristic hallmarks of neurodegenerative disorders: neuronal cell death, neuroinflammation, altered neurogenesis, and disrupted blood–brain barrier and vascular system, among others.
The clear association between neuroinflammation and Alzheimer’s and/or Parkinson’s disease led to the study of the role of CysLTs pathways and receptors in these diseases.
Alzheimer’s disease (AD, described in more detail in Section 4.1) is a neurodegenerative disease characterized by memory loss and dementia. There is evidence for CysLTR1 involvement in AD, leading to amyloidogenesis and neuroinflammation. In particular:
(1) In an AD mouse model (APP/PS1 double transgenic, overexpressing mutated forms of human amyloid precursor protein, APP, and presenilin 1), the expression of CysLTR1 was found to increase with ageing, and to correlate with Aβ deposits and behaviour deficits [84,101];
(2) LTD4 upregulates APP, β-, and γ-secretase levels, and facilitates Aβ amyloid accumulation via the CysLTR1-mediated NF-κB pathway [102,103,104].
Aggregated Aβ1–42 is known to cause AD-like neurotoxicity and cognitive deficiency, associated with pro-inflammatory cytokine production (TNF-α, IL-1β) and increased cell apoptosis [84,105,106]. Additional studies also revealed that Aβ plaques are associated with an increased oxidative stress status. Oxidative stress is known to upregulate cPLA2 activity, leading to an increased release of arachidonic acid metabolites [66]. These responses are inhibited by Lukast drugs (pranlukast, montelukast, and zafirlukast), suggesting that CysLTR1 is a pro-inflammatory regulator and is involved in AD initiation and progression [66,84,105,106].
Parkinson’s disease (PD, described in more detail in Section 4.2) is also a neurodegenerative disorder characterised by the progressive degeneration and loss of dopaminergic neurons. Inflammation induction in PD models (with rotenone or lipopolysaccharide) leads to microglia activation, increasing the production of the pro-inflammatory cytokines TNF-α, IL-1β, and IL-6, and brain inflammation, leading to dopaminergic neuronal loss [47,66,107,108,109,110]. This action was inhibited by montelukast via the CysLTR1-mediated p38 MAPK/NF-κB pathway [82,107,111], and also by selective inhibition or knockout of CysLTR2 [86], suggesting that CysLTR1 and CysLTR2 could be strategic targets against PD. CysLTR1, as well as 5-LOX, are found to be upregulated in mouse PD models [92], further strengthening the hypothesis that the LT pathway contributes to the progression of PD.
In conclusion, leukotrienes play an important role in the progression of neurodegenerative disorders. Receptors involved in the different steps of the LT cascade interfere with the inflammatory process, which is partially responsible for the development of the characteristic hallmarks of AD and PD. For this reason, targeting the CysLT pathway seems to be a promising strategy to delay the progression of these disorders.
This entry is adapted from the peer-reviewed paper 10.3390/ph15091039
This entry is offline, you can click here to edit this entry!