According to the World Health Organization (WHO), cancer is a leading cause of death worldwide, with nearly 10 million deaths in 2020 [
1]. Different conventional methods and treatments are available for cancer, such as chemotherapy, radiotherapy, and surgical resection. However, if some cancer cells escape these treatments, they can lead to more aggressive tumors [
2]; thus, these methods are insufficient. Recently, new therapies have been added to the arsenal to fight cancer with promising results, such as targeted therapy, stem cell therapy, nanoparticles, and active or passive immunotherapy [
3,
4].
The most used DNA-based vectors for cancer gene therapy and DNA vaccination are plasmids, small circular molecules originally obtained from bacteria. Furthermore plasmids, other non-plasmid DNA-based platforms for gene delivery have recently been reported. Some examples of this type of platform are minicircle DNA (the unnecessary plasmid backbone is removed by recombination) [
7], MIDGE DNA (minimalistic expression constructs) [
8], Doggybone DNA (linear, covalently closed, double-stranded molecules) [
9], or linear DNA amplicons produced by polymerase chain reaction (PCR) [
10].
2. Plasmid Design for Cancer Therapy
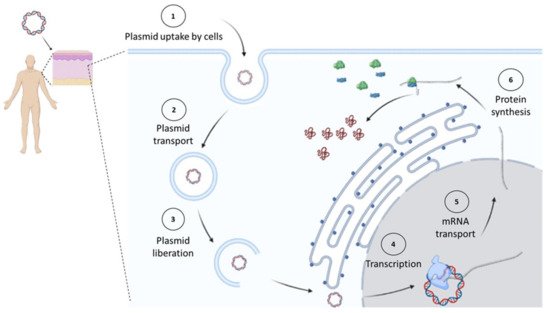
Plasmids used for cancer gene therapy or DNA vaccination must contain at least one expression cassette that directs the expression of a protein that will induce the therapeutic effect. After DNA uptake by the cell, it needs to reach the nucleus, where the gene will direct the therapeutic protein expression in the same way the cell produces its own proteins (Figure 1).
Figure 1. In vivo expression of cancer therapeutic proteins. Once a plasmid enters the cell, it must reach the nucleus, where it will start its transcription by the cell’s machinery. Later, the synthesized messenger RNA (mRNA) will be transported to the cytosol to be decoded by ribosomes into proteins. Figure created in Biorender.com.
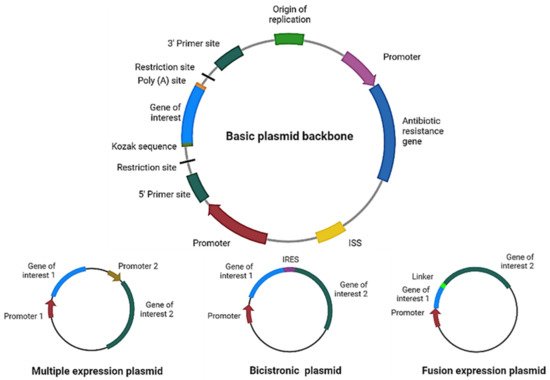
For therapy to be effective, the correct design and optimization of the plasmid are crucial (
Figure 2). For example, if more than one gene of interest needs to be expressed using a single plasmid, we can even express them independently (each gene with its own promoter), in a multicistronic system (two or more genes under the control of the same promoter), or as a fusion protein (a linker sequence between both sequences may be added). For the multicistronic system, an internal ribosome entry site (IRES) or a virus-derived T2A sequence must be placed between the different genes [
12,
13,
14,
15].
Figure 2. Plasmid design for expression of therapeutic proteins. Schematic representation of the main elements to include in a basic plasmid backbone for cancer therapy and plasmids for expression of multiple proteins. ISS: immunostimulatory sequences; IRES: internal ribosome entry site. Figure created in Biorender.com.
Codon optimization of the gene of interest is highly important, since the richness of guanines and cytosines increases messenger RNA (mRNA) levels [
16,
17]. Furthermore, the DNA molecule per se may stimulate the immune system through its unmethylated cytosine–phosphate–guanine (CpG) motifs and double-stranded structure [
18]. CpG sequences in DNA vaccines have been shown to increase immunogenicity, acting as immunostimulatory sequences (ISS) through recognition by the Toll-like receptor 9 (TLR9) present in antigen-presenting cells (APCs) [
19]; however, they may decrease gene expression [
20].
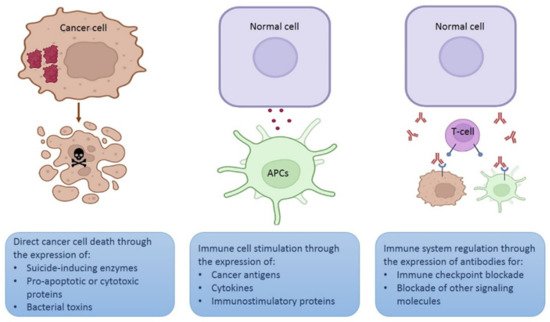
Depending on the strategy intended for the plasmid, the gene of interest may encode a therapeutic protein to kill cancer cells directly, for example, a proapoptotic protein [
21], an enzyme that activates a prodrug [
22,
23], a cytotoxic peptide [
24], or a bacterial toxin [
25,
26]. Plasmids encoding specific small interfering RNA (siRNA) molecules may be used for cancer gene therapy [
27,
28] (
Figure 3). In this case, a tumor-specific promoter can direct the transgene expression in cancer cells [
29].
Figure 3. Different strategies using plasmids for therapeutic purposes. Schematic representation of three different strategies involving in vivo expression of therapeutic proteins. Figure created in Biorender.com.
Another option is that the gene of interest encodes an antigen or a cytokine to stimulate immune cells (mainly lymphocytes or APCs) [
30,
31] to destroy cancer cells. Since a high transgene expression is required for immune stimulation, strong promoters, such as the cytomegalovirus (CMV) promoter, are more suitable for this strategy. Furthermore, expression of the therapeutic protein may be performed by any cell that captures the plasmid. In addition, APC-targeted expression may be achieved using specific promoters [
32].
A novel strategy involves using plasmids that encode monoclonal antibodies to block different signaling cascades, such as immune checkpoints or other molecules expressed on the cell surface or secreted in the tumor microenvironment [
33].
3. Tumor-Specific Promoters for Gene Therapy
As we can find cell- and tissue-specific promoters that regulate the expressions of different genes in normal cells, some promoters also allow for the expression of genes that favor the proliferation of cancer cells. Scientists have taken advantage of the nature of these to allow for the expression of therapeutic genes only in cancer cells. There are promoters functional in cancers of different origin (cancer-specific promoters) but not active in normal cells, and there are specific promoters that are active only in a limited type of cancer cells (tumor-specific promoters) [
29]. Herein, we mention some of the most widely used cancer-specific promoters, whose antitumoral effects have been analyzed in vivo using non-viral gene therapy.
The promoter of human telomerase reverse transcriptase (hTERT) has null activity in most somatic cells due to the absence of its methylation, which allows for its binding to the repressor. hTERT is a type of promoter active via methylation in different types of tumor tissues, which allows for the high expression of telomerase, an enzyme responsible for increasing telomeres in the proliferation of cancer cells [
34,
35]. The therapeutic use of this promoter in cancer therapy has been analyzed in different works. A plasmid that encodes the non-metastatic clone 23, isoform H1 (
nm23-H1) gene, a metastasis suppressor gene under the control of the hTERT promoter, inhibited tumor growth and distant metastasis when evaluated in a lung cancer xenograft model after intratumoral injection with the vector [
36]. In another work, a plasmid that encodes KK-64, a cytotoxic peptide, under the control of hTERT was administered in the form of DNA/liposome complexes to mice previously inoculated with mouse hepatocarcinoma cell line H22, with a reduction in tumor growth observed [
37]. A novel version of the hTERT promoter using a VISA (VP16-Gal4-WPRE integrated systemic amplifier) system was reported. In this work, the hTERT-VISA system was used to drive the expression of E1A, an adenoviral transcription factor with anticancer properties. Significant antitumor activity was reported in an ovarian cancer xenograft murine model after intravenous delivery of the plasmid/liposomal nanoparticles [
38].
The
BIRC5 gene is active in different cancers but not in normal tissues. It drives the expression of survivin, an apoptosis inhibitor important for cancer development [
39]. This promoter has been used in a minicircle system with potential clinical use for prostate cancer diagnosis and treatment [
40]. In another work, the survivin promoter was used in combination with hTERT promoter to form a hybrid promoter to increase its strength of expression in transfected cancer cells. This hybrid promoter directed the expression of Herpes simplex virus-1 thymidine kinase (HSVtk) and the mouse granulocyte-macrophage colony-stimulating factor (GM-CSF). These transfected cancer cells were implanted in mice, and tumor growth inhibition was observed [
41].
A candidate promoter for breast cancer is Erb-B2 receptor tyrosine kinase 2 (
ERBB2) gene promoter; however, this is expressed in only 20–25% of tumors [
42,
43,
44], and it is also active in prostate, pancreas, colon, and ovary cancer cells [
45,
46,
47]. The
ERBB2 gene promoter has been used in some works, as in a clinical trial for breast cancer where the patients received intratumoral injection of a plasmid that encodes the
E. coli cytosine deaminase under the control of the
ERBB2 gene promoter to activate the prodrug fluorocytosine [
48]. In another work, a plasmid containing a minimum version of this promoter directing the expression of HSVtk to confer selective cytotoxicity to ganciclovir was constructed and proved in nude mice bearing human breast cancer cells. The administration of ganciclovir in human breast cancer cells transfected with this plasmid reduced tumor growth [
49].
Regarding lung cancer, the thyroid transcription factor-1 (TTF-1) promoter is active in small cell lung carcinoma and adenocarcinoma [
50,
51]. Low constitutive expression is found in healthy lung cells, such as type II alveolar cells [
52]. The use of this promoter to drive the expression of miR-7, a powerful tumor suppressor, was reported. This study showed the targeting of transgene expression in the tumor cells via a remote hypodermic injection of a plasmid, downregulating tumor growth in a nude mice model of lung cancer [
53].
Prostate-specific antigen (PSA) is regulated by the prostate cancer promoter, which has low constitutive expression in the prostate epithelium [
54]; however, high levels are detected in patients with metastatic prostate cancer [
55]. It is known that the activity of this promoter can be regulated by DNA-binding proteins [
55], and this regulation may be androgen dependent or independent [
54,
56]. A recently published work reported using liposomes with a vector containing the PSA promoter driving the expression of perforin (a protein that makes pores on the plasma membrane) in cancer cells. After intravenous administration of this therapy, a reduced tumor volume was observed in a xenograft model of prostate cancer [
57].