Your browser does not fully support modern features. Please upgrade for a smoother experience.
Please note this is an old version of this entry, which may differ significantly from the current revision.
Obesity incidence is rising worldwide, including women of reproductive age, contributing to increased gestations in which Maternal Obesity (MO) occurs. Offspring born to obese mothers present an increased predisposition to develop metabolic (e.g., obesity, diabetes) and cardiovascular disease (CVD). The developmental programming of the metabolic dysfunction in MO offspring can initiate in utero. The different availability of metabolic substrates, namely glucose, can modulate cellular growth, proliferation, and differentiation, resulting in different levels of tissue maturation and function.
- maternal overweight
- endocrine dysregulation
- metabolic dysfunction
1. Introduction
Obesity has been rapidly increasing worldwide, especially among women of childbearing age. In Europe, it is estimated that 26.8–54% of reproductive-aged women are either overweight or obese [1]. More than an imbalance between energy intake and expenditure, obesity results from the interaction between genetic, behavioral, endocrine, physiologic, and other factors [2].
Being overweight or obese before pregnancy represents an increased risk for excessive gestational weight gain (GWG), whose prevalence reaches up to 40% of pregnancies [3]. Maternal obesity (MO) includes a high pre-gestational body mass index and/or excessive GWG. MO is associated with short- and long-term adverse outcomes for both the mother and the offspring [4]. In the mothers, MO is related to increased odd-ratios for the long-term development of obesity, metabolic syndrome, type 2 diabetes mellitus (T2DM), and cardiovascular disease (CVD) [5]. During pregnancy, MO is associated with a higher risk of developing pregnancy-related disorders such as gestational diabetes mellitus (GDM), hypertensive disorders (e.g., gestational hypertension, pre-eclampsia), venous thromboembolism, and wound infection [6][7][8][9][10]. MO is also associated with increased risks of complications during delivery: cesarean delivery risk is increased 1.41-fold in overweight and 1.75-fold in obese women, usually associated with protracted labor and cephalopelvic disproportion that result from fetal macrosomia [11]. In addition, fetuses of overweight women present an elevated risk of incidence of structural birth defects(e.g., neural tube defects), prematurity, macrosomia, hypoglycemia, birth injury from shoulder dystocia, and stillbirth [6].
According to the Developmental Origins of Health and Disease (DOHaD) hypothesis, in response to the intrauterine environment, a single genotype may give origin to an array of fetal physiological and/or morphological state alterations [12]. It is now accepted that maternal lifestyle before and during pregnancy influences lifelong offspring’s metabolism and responses to metabolic demands, predisposing them to non-communicable diseases (NCDs), including obesity, T2DM, and CVD [13]. Offspring’s early consequences of MO generally include fetal macrosomia. MO offspring can present low gestational weight instead if MO is concomitant with hypertension and compromised placental vasculature. Offspring from MO also have a higher occurrence of obesity and diabetes in childhood as well as CVD in young adulthood [14][15][16]. The clinical evidence reports that newborns delivered by obese mothers present a three-fold increased risk of developing obesity [3], a higher tendency to be large for gestational age (LGA) [17], an adverse lipid profile [3], increased body-adiposity [18], and an abnormal body fat mass distribution [1]. Moreover, adults born to obese mothers present an increased body mass index (BMI) in comparison with adults born to lean mothers in Australian [19], Israeli [20], and Finnish [21] populations. Offspring born to mothers with excessive GWG present increased adiposity, especially for higher GWG at early pregnancy, and increased high-density lipoprotein, triglycerides, and inflammatory biomarkers in mid-pregnancy excessive GWG [18]. Clinical studies have reported sexual dimorphism in response to MO. MO-offspring 6-year-old males presented greater adiposity than males born to lean mothers, whereas no differences for females were detected [22].
These data highlight that, beyond genetic predisposition and postnatal environment, NCDs transmission can be intergenerationally (direct impact on first-generation health or impact on the second generation through the programming of first-generation germline cells) or even transgenerationally programmed (vicious cycle of obesity transmission across generations without genetic explanation) [23]. To prevent the vicious cycle of transmission among generations and provide more specialized prenatal and postnatal care, it is necessary to understand the molecular mechanisms underlying the development of NCDs in MO offspring.
2. Mechanistic Links between MO and the Development of Metabolic Disease in the Offspring
The evidence in clinical studies of increased disease incidence in MO offspring has made it necessary to explore the mechanistic links between offspring metabolic disease development and MO. For ethical reasons, animal studies are critical to examining the cellular and molecular mechanisms associated with the phenotypes of the human offspring born to MO. Several animal models have been used in an attempt to elucidate these mechanisms. Studies in rats, sheep, and non-human primates have consistently proven MO’s deleterious effects on offspring, displaying similar phenotypes to those described in humans [24]. Increased birth- and postnatal-body weight [25][26][27], excessive body fat percentage [28][29], adiposity [29][30] and hyperinsulinemia [28][30] are frequent in rodent, ovine, and non-human primate studies ranging from MO offspring’s early ages until adulthood. These outcomes are often accompanied by an abnormal lipidic profile (increased cholesterol and triglycerides levels) [28] and endocrine dysregulation, evidenced by impaired adipokine levels [15][21][31].
The mechanisms associated with these outcomes and, subsequently, increased predisposition to metabolic disease for MO progeny seem to be multifactorial. In this focused manuscript, researchers explore (a) metabolic programming and oxidative stress, (b) epigenomic alterations, and (c) endocrine-stress response dysregulation as the main potential mechanisms involved in offspring’s metabolic disease programming by MO. Researchers define the first two as the first hit of offspring adaptations to MO, i.e., structural and molecular alterations already observed at fetal developmental stages that are maintained throughout offspring life. The first hit of developmental programming is tissue-specific and a direct result of the intrauterine environment on each organ cell’s maturation, growth, and proliferation.
MO influences fetal nutrient availability, namely glucose and lipids, due to the different blood maternal metabolome and the ability of each molecule to cross through the placenta [32]. For example, glucose is only partially buffered by the placenta, meaning that if a higher glucose concentration exists in maternal blood, it will also be increased in fetal circulation [33]. This leads to fetal hyperinsulinemia, which potentiates insulin resistance, metabolic dysregulation, and oxidative stress; these are thought to be involved in offspring’s metabolic disease programming [16]. Accordingly, insulin resistance in white adipose, hepatic, and cardiac tissues of MO offspring has been reported [16]. The hepatic and cardiac ratio between phosphorylated insulin receptor substrate 1 (IRS-1) at Ser307 residue and total IRS-1 is increased for 3- [34] and 4-month-old [26] MO mice offspring. In addition, many studies have shown mitochondrial dysfunction in individuals with insulin resistance (IR) [35]. MO offspring present sex-specific cardiac mitochondrial function modulation [36]. Newborn female MO rat offspring present increased expression of mitochondrial fusion proteins, and males show increased levels of proteins involved in mitophagy [36]. Four-month-old MO mice offspring present altered cardiac mitochondrial ultrastructure, with swollen and disrupted mitochondria, reduced mitochondrial membrane potential and increased ROS production [26]. Cardiac oxidative stress was determined by higher malondialdehyde levels in newborn rat MO offspring [37]. Hepatic oxidative stress in non-human primates [31] and 20-week-old male mice MO offspring [38] was marked by increased levels of 8-hydroxy-2-deoxyguanosine. Increased pancreatic glutathione peroxidase mRNA levels were detected in 20-week-old male mice MO offspring [38], potentially as a way to counteract oxidative stress.
Although previously hypothesized, it remains unknown whether MO-induced metabolic programming in the offspring has an effect on the long-term regulation of gene expression [16]. In spite of this, the literature has pointed to epigenetic dysregulation as a possible disease-programming mechanism due to the fetal epigenome responsiveness to maternal dietary patterns [39]. MO-induced microRNA (miRNA) expression alterations, DNA methylation, and post-translational histone modifications are the main mechanisms that lead to the epigenetic modulation of gene expression in offspring [40]. Hepatic miRNAs that regulate the insulin-signaling pathway are decreased in 4-month-old MO sheep offspring [39]. The inhibition of IRS-1 translation in white adipose tissue (WAT) is provoked by elevated miRNA-126 in 2-month-old MO mice offspring [41]. Genome-scale DNA methylation is altered by MO in 21-day-old rat male offspring [42]. Differential DNA methylation patterns and post-translational histone modifications (acetylation/methylation) were also found for genes encoding for adipocytokine, adiponectin, and leptin in MO mice offspring [43].
The programming of hypothalamic appetite-regulating centers is another mechanism potentially involved in MO programming of offspring’s metabolic disease [44]. MO induces endocrine stress-response dysregulation, leading to offspring’s hyperphagia and obesity [16]. The orexigenic drive (i.e., increased food intake, weight gain, the ratio between orexigenic and anorexigenic neuronal number/peptide expression and signaling) is increased in MO offspring [45]. Some studies have suggested that, in rodents, the regulation of the central nervous system (CNS) occurs postnatally; however, recent research has indicated that this might occur in utero [43]. In the hypothalamus of newborn MO rat offspring, neuropeptide Y mRNA expression and protein levels are increased [45]. In contrast, in another study, 1-day-old MO offspring’s hypothalamus presented decreased mRNA expression of the leptin receptor, signal transducer/activator of transcription (STAT)-3, and hypothalamic neuropeptide Y [46], demonstrating that appetite-regulating centers are likely modulated by the MO-induced intrauterine environment. Nevertheless, current research fails to explain this contradictory data. Further research is needed to unravel this issue and whether MO-induced endocrine dysregulation in the hypothalamus extends to other organs and remains in adulthood.
In addition to biochemical end-point alterations, evidence supports deeper alterations, with the fetal tissues and organs adapting their function to an adverse intrauterine environment caused by MO [47], leading to organ dysfunction that could persist over the offspring’s life course and lead to metabolic disease development (Figure 1). For a detailed review of the mechanisms associated with the first hit of developmental programming by MO, consult the reviews [16][35].
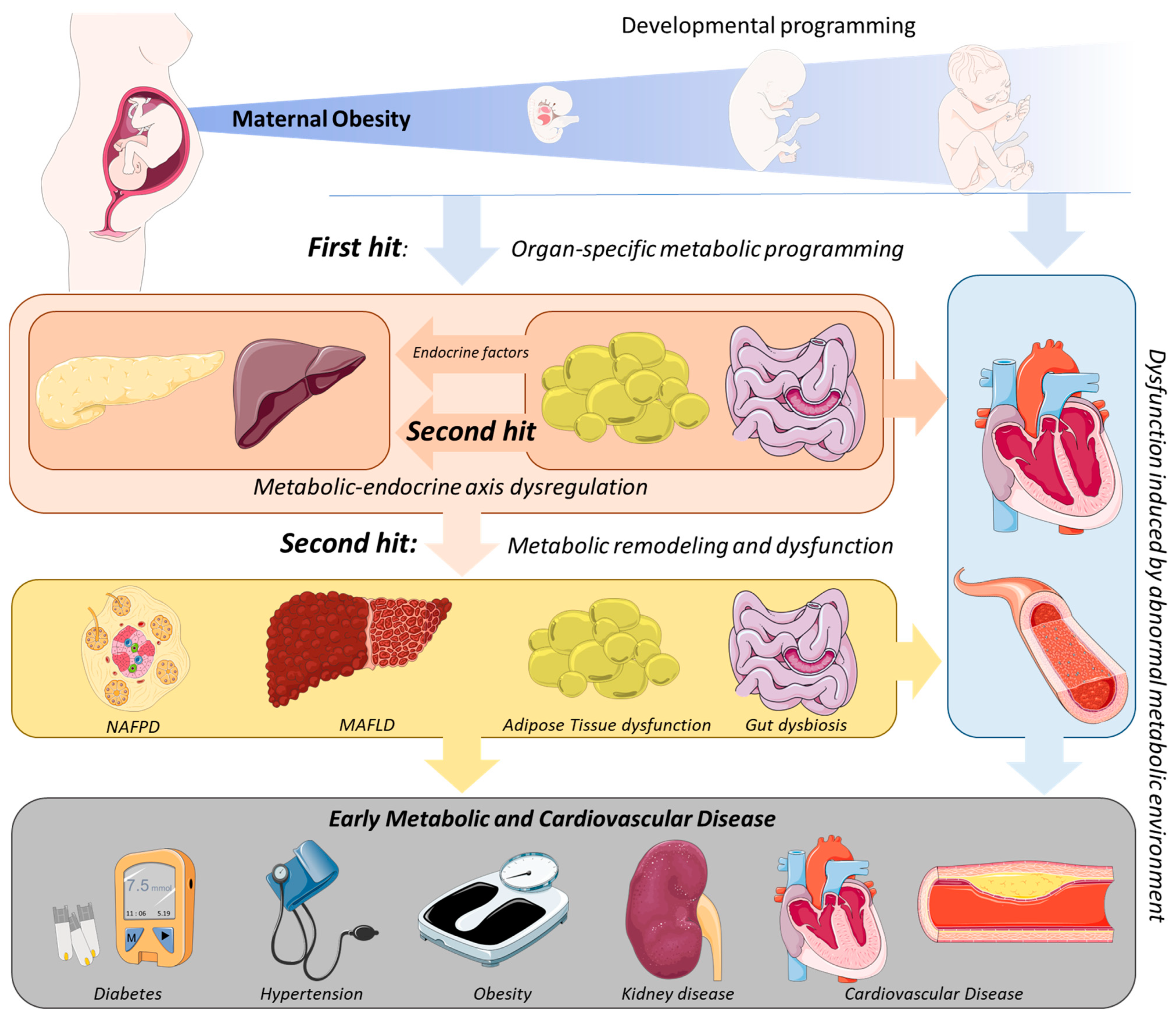
Figure 1. Mechanism of endocrine-metabolic axis programming in offspring due to maternal obesity. Throughout fetal development, Maternal Obesity (MO) programs organ-specific dysfunction through the modulation of cell proliferation, growth rates, differentiation, and maturation—the first hit of developmental programming. Consequently, metabolic dysregulation is commonly observed in organs in early life stages, which leads to different metabolic homeostasis in MO offspring. Among these, the adipose tissue and gut are organs with an important endocrine role in the endocrine–metabolic axis. Adipose tissue dysfunction and gut dysbiosis are early organ-specific events in MO offspring, promoting the release of endocrine proteins (e.g., adipokines, glucagon-like peptide (GLP) 1/2) capable of modulating other tissues’ functions, such as the liver and pancreas, and through them, the body’s metabolism—the second hit of the endocrine–metabolic axis programming. These events can modulate the concentrations and classes of circulating metabolites (e.g., peptides, enzymes, fatty acids), which affect other organs and predispose them to metabolic diseases at early life stages.
This entry is adapted from the peer-reviewed paper 10.3390/obesities2030019
References
- González-Plaza, E.; Bellart, J.; Martínez-Verdú, M.; Arranz, A.; Luján-Barroso, L.; Seguranyes, G. Prevalencia de sobrepeso y obesidad preconcepcional en mujeres gestantes, y relación con los resultados maternos y perinatales. Enfermería Clínica 2021, 32, S23–S30.
- Unamuno, X.; Gómez-Ambrosi, J.; Rodríguez, A.; Becerril, S.; Frühbeck, G.; Catalán, V. Adipokine dysregulation and adipose tissue inflammation in human obesity. Eur. J. Clin. Investig. 2018, 48, e12997.
- Gaillard, R.; Felix, J.; Duijts, L.; Jaddoe, V.W. Childhood consequences of maternal obesity and excessive weight gain during pregnancy. Acta Obstet. Gynecol. Scand. 2014, 93, 1085–1089.
- Catalano, P.M.; Shankar, K. Obesity and pregnancy: Mechanisms of short term and long term adverse consequences for mother and child. BMJ 2017, 356, j1.
- Mouzon, S.H.-D.; Lassance, L. Endocrine and metabolic adaptations to pregnancy; impact of obesity. Horm. Mol. Biol. Clin. Investig. 2015, 24, 65–72.
- Kriebs, J.M. Obesity in pregnancy—Addressing risks to improve outcomes. J. Perinat. Neonatal Nurs. 2014, 28, 32–40.
- Bodnar, L.M.; Catov, J.M.; Klebanoff, M.A.; Ness, R.B.; Roberts, J.M. Prepregnancy Body Mass Index and the Occurrence of Severe Hypertensive Disorders of Pregnancy. Epidemiology 2007, 18, 234–239.
- Torloni, M.R.; Betrán, A.P.; Horta, B.L.; Nakamura, M.U.; Atallah, A.N.; Moron, A.F.; Valente, O. Prepregnancy BMI and the risk of gestational diabetes: A systematic review of the literature with meta-analysis: Diagnostic in Obesity and Complications. Obes. Rev. 2009, 10, 194–203.
- Callaghan, W.M.; Chu, S.Y.; Kim, Y.S.; Schmid, C.H.; Lau, J.; England, J.L.; Dietz, M.P. Maternal obesity and risk of gestational. Diabetes Care 2007, 30, 2070–2076.
- Solomon, C.G. A prospective study of pregravid determinants of gestational diabetes mellitus. J. Am. Med. Assoc. 1997, 278, 1078.
- Chu, S.Y.; Kim, S.Y.; Schmid, C.H.; Dietz, P.M.; Callaghan, W.M.; Lau, J.; Curtis, K.M. Maternal obesity and risk of cesarean delivery: A meta-analysis. Obes. Rev. 2007, 8, 385–394.
- Nijland, M.; Ford, S.P.; Nathanielsz, P. Prenatal origins of adult disease. Curr. Opin. Obstet. Gynecol. 2008, 20, 132–138.
- Godfrey, K.M.; Reynolds, R.M.; Prescott, S.L.; Nyirenda, M.; Jaddoe, V.W.V.; Eriksson, J.G.; Broekman, B.F.P. Influence of maternal obesity on the long-term health of offspring. Lancet Diabetes Endocrinol. 2016, 5, 53–64.
- Howell, K.; Powell, T.L. Effects of maternal obesity on placental function and fetal development. Reproduction 2017, 153, R97–R108.
- Yu, Y.; Arah, O.A.; Liew, Z.; Cnattingius, S.; Olsen, J.; Sorensen, H.T.; Qin, G.; Li, J. Maternal diabetes during pregnancy and early onset of cardiovascular disease in offspring: Population based cohort study with 40 years of follow-up. BMJ 2019, 367, l6398.
- Grilo, L.F.; Tocantins, C.; Diniz, M.S.; Gomes, R.M.; Oliveira, P.J.; Matafome, P.; Pereira, S.P. Metabolic Disease Programming: From Mitochondria to Epigenetics, Glucocorticoid Signalling and Beyond. Eur. J. Clin. Investig. 2021, 51, e13625.
- Yu, Z.; Han, S.; Zhu, J.; Sun, X.; Ji, C.; Guo, X. Pre-Pregnancy Body Mass Index in Relation to Infant Birth Weight and Offspring Overweight/Obesity: A Systematic Review and Meta-Analysis. PLoS ONE 2013, 8, e61627.
- Gaillard, R.; Steegers, E.A.P.; Franco, O.; Hofman, A.; Jaddoe, V.W.V. Maternal weight gain in different periods of pregnancy and childhood cardio-metabolic outcomes. The Generation R Study. Int. J. Obes. 2014, 39, 677–685.
- Mamun, A.A.; O’Callaghan, M.; Callaway, L.; Williams, G.; Najman, J.; Lawlor, D.A. Associations of gestational weight gain with offspring body mass index and blood pressure at 21 years of age: Evidence from a birth cohort study. Circulation 2009, 119, 1720–1727.
- Hochner, H.; Friedlander, Y.; Calderon-Margalit, R.; Meiner, V.; Sagy, Y.; Avgil-Tsadok, M.; Burger, A.; Savitsky, B.; Siscovick, D.S.; Manor, O. Associations of maternal prepregnancy body mass index and gestational weight gain with adult offspring cardiometabolic risk factors: The Jerusalem Perinatal Family Follow-up Study. Circulation 2012, 125, 1381–1389.
- Eriksson, J.; Sandboge, S.; Salonen, M.; Kajantie, E.; Osmond, C. Maternal weight in pregnancy and offspring body composition in late adulthood: Findings from the Helsinki Birth Cohort Study (HBCS). Ann. Med. 2015, 47, 94–99.
- Andres, A.; Hull, H.R.; Shankar, K.; Casey, P.H.; Cleves, M.A.; Badger, T.M. Longitudinal body composition of children born to mothers with normal weight, overweight, and obesity. Obesity 2015, 23, 1252–1258.
- Bronson, S.C.; Seshiah, V. Transgenerational transmission of non-communicable diseases: How to break the vicious cycle? Cureus 2021, 13, 18754.
- Menting, M.D.; Mintjens, S.; van de Beek, C.; Frick, C.J.; Ozanne, S.E.; Limpens, J.; Roseboom, T.J.; Hooijmans, C.R.; van Deutekom, A.W.; Painter, R.C. Maternal obesity in pregnancy impacts offspring cardiometabolic health: Systematic review and meta-analysis of animal studies. Obes. Rev. 2019, 20, 675–685.
- Nunes, F.; Frantz, E.D.C.; Lannes, W.R.; Menezes, M.C.D.S.; Mandarim-De-Lacerda, C.; Souza-Mello, V. Pregestational maternal obesity impairs endocrine pancreas in male F1 and F2 progeny. Nutrition 2015, 31, 380–387.
- Turdi, S.; Ge, W.; Hu, N.; Bradley, K.M.; Wang, X.; Ren, J. Interaction between maternal and postnatal high fat diet leads to a greater risk of myocardial dysfunction in offspring via enhanced lipotoxicity, IRS-1 serine phosphorylation and mitochondrial defects. J. Mol. Cell. Cardiol. 2012, 55, 117–129.
- Rivera, H.M.; Kievit, P.; Kirigiti, M.A.; Bauman, L.A.; Baquero, K.; Blundell, P.; Dean, T.A.; Valleau, J.C.; Takahashi, D.L.; Frazee, T.; et al. Maternal high-fat diet and obesity impact palatable food intake and dopamine signaling in nonhuman primate offspring. Obesity 2015, 23, 2157–2164.
- Rajia, S.; Chen, H.; Morris, M.J. Maternal overnutrition impacts offspring adiposity and brain appetite markers-modulation by postweaning diet. J. Neuroendocr. 2010, 22, 905–914.
- Long, N.M.; George, L.A.; Uthlaut, A.B.; Smith, D.T.; Nijland, M.; Nathanielsz, P.; Ford, S.P. Maternal obesity and increased nutrient intake before and during gestation in the ewe results in altered growth, adiposity, and glucose tolerance in adult offspring1. J. Anim. Sci. 2010, 88, 3546–3553.
- Chen, H.; Simar, D.; Morris, M.J. Maternal obesity impairs brain glucose metabolism and neural response to hyperglycemia in male rat offspring. J. Neurochem. 2013, 129, 297–303.
- McCurdy, C.; Bishop, J.M.; Williams, S.M.; Grayson, B.E.; Smith, M.S.; Friedman, J.E.; Grove, K.L. Maternal high-fat diet triggers lipotoxicity in the fetal livers of nonhuman primates. J. Clin. Investig. 2009, 119, 323–335.
- Hirschmugl, B.; Perazzolo, S.; Sengers, B.G.; Lewis, R.M.; Gruber, M.; Desoye, G.; Wadsack, C. Placental mobilization of free fatty acids contributes to altered materno-fetal transfer in obesity. Int. J. Obes. 2021, 45, 1114–1123.
- Hay, W.W. Placental-fetal glucose exchange and fetal glucose metabolism. Trans. Am. Clin. Clim. Assoc. 2006, 117, 321–340.
- Martin-Gronert, M.S.; Fernandez-Twinn, D.S.; Poston, L.; Ozanne, S.E. Altered hepatic insulin signalling in male offspring of obese mice. J. Dev. Orig. Heal. Dis. 2010, 1, 184–191.
- Tocantins, C.; Diniz, M.S.; Grilo, L.F.; Pereira, S.P. The birth of cardiac disease: Mechanisms linking gestational diabetes mellitus and early onset of cardiovascular disease in offspring. WIREs Mech. Dis. 2022.
- Larsen, T.D.; Sabey, K.H.; Knutson, A.J.; Gandy, T.C.T.; Louwagie, E.J.; Lauterboeck, L.; Mdaki, K.S.; Baack, M.L. Diabetic Pregnancy and Maternal High-Fat Diet Impair Mitochondrial Dynamism in the Developing Fetal Rat Heart by Sex-Specific Mechanisms. Int. J. Mol. Sci. 2019, 20, 3090.
- Mdaki, K.S.; Larsen, T.D.; Wachal, A.L.; Schimelpfenig, M.D.; Weaver, L.J.; Dooyema, S.D.R.; Louwagie, E.J.; Baack, M.L. Maternal high-fat diet impairs cardiac function in offspring of diabetic pregnancy through metabolic stress and mitochondrial dysfunction. Am. J. Physiol. Circ. Physiol. 2016, 310, H681–H692.
- Yokomizo, H.; Inoguchi, T.; Sonoda, N.; Sakaki, Y.; Maeda, Y.; Inoue, T.; Hirata, E.; Takei, R.; Ikeda, N.; Fujii, M.; et al. Maternal high-fat diet induces insulin resistance and deterioration of pancreatic β-cell function in adult offspring with sex differences in mice. Am. J. Physiol. Metab. 2014, 306, E1163–E1175.
- Nicholas, L.M.; Rattanatray, L.; MacLaughlin, S.M.; Ozanne, S.E.; Kleemann, D.O.; Walker, S.K.; Morrison, J.L.; Zhang, S.; Muhlhäusler, B.S.; Martin-Gronert, M.S.; et al. Differential effects of maternal obesity and weight loss in the periconceptional period on the epigenetic regulation of hepatic insulin-signaling pathways in the offspring. FASEB J. 2013, 27, 3786–3796.
- Agarwal, P.; Morriseau, T.S.; Kereliuk, S.M.; Doucette, C.A.; Wicklow, B.A.; Dolinsky, V.W. Maternal obesity, diabetes during pregnancy and epigenetic mechanisms that influence the developmental origins of cardiometabolic disease in the offspring. Crit. Rev. Clin. Lab. Sci. 2018, 55, 71–101.
- Fernandez-Twinn, D.S.; Alfaradhi, M.Z.; Martin-Gronert, M.S.; Duque-Guimaraes, D.E.; Piekarz, A.; Ferland-McCollough, D.; Bushell, M.; Ozanne, S.E. Downregulation of IRS-1 in adipose tissue of offspring of obese mice is programmed cell-autonomously through post-transcriptional mechanisms. Mol. Metab. 2014, 3, 325–333.
- Borengasser, S.J.; Zhong, Y.; Kang, P.; Lindsey, F.; Ronis, M.J.J.; Badger, T.M.; Gomez-Acevedo, H.; Shankar, K. Maternal Obesity Enhances White Adipose Tissue Differentiation and Alters Genome-Scale DNA Methylation in Male Rat Offspring. Endocrinology 2013, 154, 4113–4125.
- Masuyama, H.; Hiramatsu, Y. Effects of a High-Fat Diet Exposure in UTERO on the Metabolic Syndrome-Like Phenomenon in Mouse Offspring through Epigenetic Changes in Adipocytokine Gene Expression. Endocrinology 2012, 153, 2823–2830.
- Zambrano, E.; Ibáñez, C.; Martínez-Samayoa, P.M.; Lomas, C.; Durand-Carbajal, M.; González, G.L.R. Maternal Obesity: Lifelong Metabolic Outcomes for Offspring from Poor Developmental Trajectories During the Perinatal Period. Arch. Med. Res. 2016, 47, 1–12.
- Desai, M.; Ross, M.G. Maternal-infant nutrition and development programming of offspring appetite and obesity. Nutr. Rev. 2020, 78, 25–31.
- Morris, M.J.; Chen, H. Established maternal obesity in the rat reprograms hypothalamic appetite regulators and leptin signaling at birth. Int. J. Obes. 2008, 33, 115–122.
- Shrestha, N.; Ezechukwu, H.C.; Holland, O.J.; Hryciw, D.H. Developmental programming of peripheral diseases in offspring exposed to maternal obesity during pregnancy. Am. J. Physiol. Integr. Comp. Physiol. 2020, 319, R507–R516.
This entry is offline, you can click here to edit this entry!