Australia produces high-quality sweet cherries and generates revenue from local and export markets. Due to increased demand in the markets, the area of sweet cherry production has increased in Australia. Sweet cherry breeding and production have challenges such as self-incompatibility genotypes and phenotyping of agronomic, physiological, and quality traits. Phenotyping is identified as an interaction between cultivars and plant responses to different environmental conditions which determine the plant yield and fruit quality. Efficient phenotyping techniques are important to develop new sweet cherry cultivars in Australia with high yield potential, high fruit quality, temperature tolerance (cold and heat), and drought tolerance.
- environmental factors
- fruiting spur
- fruit development
1. Phenotyping Based on Agronomic and Physiological Traits in Sweet Cherries
1.1. Agronomic Traits
1.2. Physiological Traits
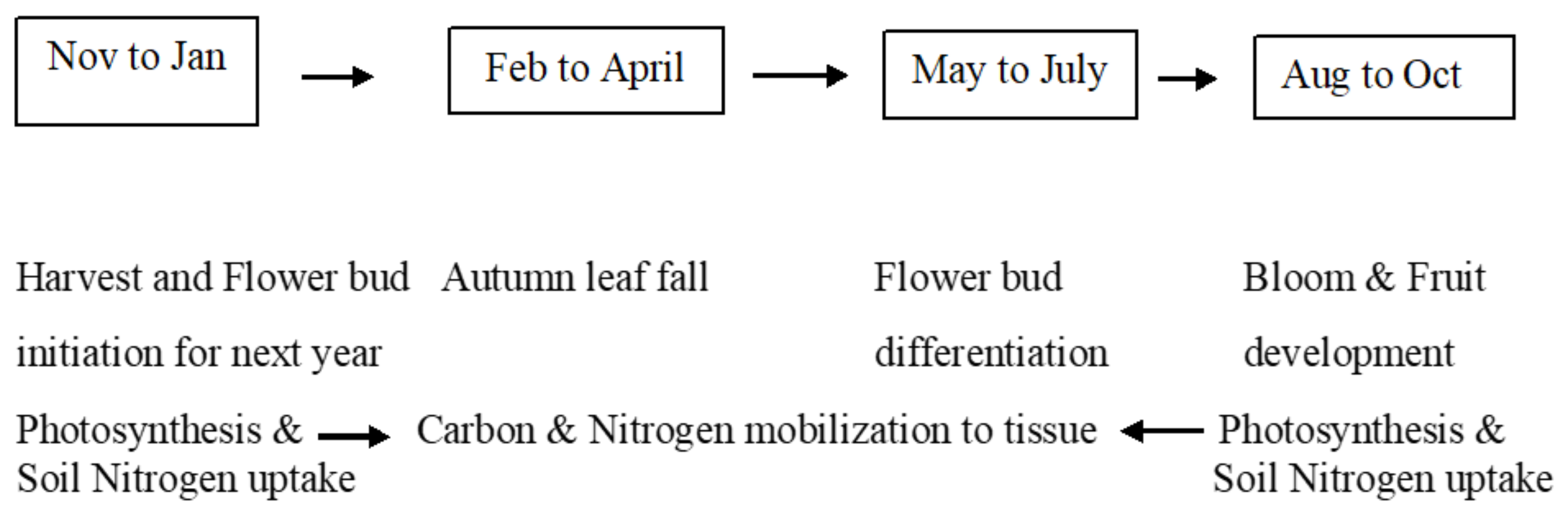
1.3. Impact of Environmental Factors in Agronomic, Physiological and Reproductive Traits
2. Phenotyping Based on Fruit Quality Traits in Sweet Cherries

| Firmness | g/m2 | Sweetness | Brix % |
|---|---|---|---|
| Hard | >350 | Super sweet | ≥21 |
| Very firm | 300–350 | Very sweet | 19–20 |
| Firm | 240–300 | Sweet | 17–18 |
| Less firm | <240 | Moderate sweet | 16 |
This entry is adapted from the peer-reviewed paper 10.3390/horticulturae8080694
References
- Hardner, C.M.; Hayes, B.J.; Kumar, S.; Vanderzande, S.; Cai, L.; Piaskowski, J.; Quero-Garcia, J.; Campoy, J.A.; Barreneche, T.; Giovannini, D.; et al. Prediction of genetic values of sweet cherry fruit maturity among environments using a 6K SNP array. Hortic. Res. 2019, 6, 6.
- Yue, C.R.; Gallardo, K.; Luby, J.J.; Rihn, A.L.; McFerson, J.R.; McCracken, V.; Oraguzie, N.; Weebadde, C.; Sebolt, A.; Iezzoni, A. An evaluation of U.S. tart and sweet cherry producer’s trait prioritization: Evidence from audience surveys. HortScience 2014, 49, 931–937.
- Quero-Garcia, J.; Fodor, A.; Reignier, A.; Capdeville, G.; Joly, J.; Tauzin, Y.; Fouilhaux, L.; Direlwanger, E. QTL detection of important agronomic traits for sweet cherry production. Acta Hortic. 2014, 1020, 57–64.
- Etienne, C.; Rothan, C.; Moing, A.; Plomion, C. Candidate genes and QTLs for sugar and organic acid content in peach . Theor. Appl. Genet. 2002, 105, 145–159.
- Salazar, J.A.; Ruiz, D.; Campoy, J.A.; Tartarini, S.; Dondini, L.; Martinez-Gomez, P. Inheritance of reproductive phenology traits and related QTL identification in apricot. Tree Genet. Genomes 2016, 12, 71.
- Tukey, H.B. Growth of the peach embryo in relation to growth of fruit and season of ripening. Proc. Amer. Soc. Hort. Sci. 1933, 30, 208–218.
- Tukey, H.B. Embryo abortion in early-ripening varieties of Prunus avium. Bot. Gaz. 1933, 94, 433–468.
- Braak, J.P. The effect of flowering date and temperature on embryo development in sweet cherry Prunus avium L. Neth. J. Agric. Sei. 1978, 26, 13–30.
- Marti, A.F.; Athanson, B.; Koepke, T.; Font-Forcada, C.; Dhingra, A.; Oraguzie, N. Genetic diversity and relatedness of sweet cherry (Prunus avium L.) cultivars based on single nucleotide polymorphic markers. Front. Plant Sci. 2012, 3, 116.
- Azarenko, A.N.; Chozinski, A.; Brewer, L.J. Fruit growth curve analysis of seven sweet cherry cultivars. Acta Hortic. 2008, 795, 561–565.
- Brunt, C. Cherry Cultivar Selection: Chill Hours and Climate Change; Horticulture Australia Ltd.: North Sydney, Australia, 2012.
- Prasad, P.V.V.; Djanaguiraman, M. Response of floret fertility and individual grain weight of wheat to high temperature stress: Sensitive stages and thresholds for temperature and duration. Funct. Plant Biol. 2014, 41, 1261–1269.
- Devasirvatham, V.; Gaur, P.M.; Mallikarjuna, N.; Raju, T.N.; Trethowan, R.M.; Tan, D.K.Y. Effect of high temperature on the reproductive development of chickpea genotypes under controlled environments. Funct. Plant Biol. 2012, 39, 1009–1018.
- Prasad, P.V.V.; Craufurd, P.Q.; Summerfield, R.J. Sensitivity of peanut to timing of heat stress during reproductive development. Crop Sci. 1999, 39, 1352–1359.
- James, P. Australian Cherry Production guide. Cherry Growers of Australia Inc. 2011. Available online: www.cherrygrowers.org.au (accessed on 28 January 2022).
- Dordevic, B.; Durovic, D.B.; Zec, G.N.; Boscov, D.D. The influence of rootstocks on the sensitivity of flower buds to frost and the main properties of the Carmen sweetcherry cultivar. J. Agri. Sci. 2021, 66, 351–358.
- Dziedzic, E.; Bieniasz, M.; Kowalczyk, B. Morphological and physiological features of sweet cherry floral organ affecting the potential fruit crop in relation to the rootstock. Sci. Hortic. 2019, 251, 127–135.
- Baji, M.E.; Hafida, H.; En-Nahli, S.; Company, R.S.; Kodad, O. Morphological and Pomological Characteristics of Sweet Cherry (Prunus avium L.) Grown In-situ under South Mediterranean Climate in Morocco. Int. J. Fruit Sci. 2021, 21, 52–65.
- Rakonjac, V.; Mratinic, E.; Jovkovic, R.; Aksic, F.M. Analysis of morphological variability in wild cherry (Prunus avium L.) genetic resources from Central Serbia. J. Agric. Sci. Tech. 2014, 16, 151–162.
- Ganopoulos, I.; Moysiadis, T.A.; Xanthopoulou, M.; Ganopoulou, E.; Avramidou, F.A.; Aravanopoulos, E.; Tani, P.; Madesis, A.; Tsaftaris, A.; Kazantzis, K. Diversity of morpho-physiological traits in worldwide sweet cherry cultivars of Gene Bank collection using multivariate analysis. Sci. Hortic. 2015, 197, 381–391.
- Bondarenko, P. Physiological basis of sweet cherry productivity depending on rootstocks, interstems and plant density. Open Agric. 2019, 6, 267–274.
- Roper, T.R.; Keller, J.D.; Loescher, W.H.; Rom, C.R. Photosynthesis and carbohydrate partitioning in sweet cherry: Fruiting effects. Physi. Plan. 2006, 71, 42–47.
- Demirsoy, H.; Demirsoy, L. A study on the relationships between some fruit characteristics in cherries. Fruits 2004, 59, 219–223.
- Kappel, F.; Fisher-Fleming, B.; Hogue, E. Fruit characteristics and sensory attributes of an ideal sweet cherry. HortScience 1996, 31, 443–446.
- Ayala, M.; Lang, G.A. 13C-Photoassimilate partitioning in sweet cherry (Prunus avium) during early spring. Sci. Agr. Res. 2015, 42, 191–203.
- Ayala, M.; Lang, G.A. Examining the influence of different leaf populations in sweet cherry fruit quality. Acta Hortic. 2004, 636, 481–488.
- Ayala, M.; Mora, L.; Torreblanca, J. Effect of pre-bloom pruning on 13C and 15N distribution during early spring in sweet cherry. HortScience 2018, 53, 805–809.
- Macit, I.; Lang, G.A.; DemIrsoy, H. Bud management affects fruit wood, growth, and precocity of cherry trees. Turk. J. Agr. For. 2017, 41, 42–49.
- Goncalves, B.; Moutinho-Pereira, J.; Santos, A.; Silva, A.P.; Bacelar, E.; Correia, C.; Rosa, E. Scion-rootstock interaction affects the physiology and fruit quality of sweet cherry. Tree Physiol. 2006, 26, 93–104.
- Li, B.; Xie, Z.; Zhang, A.; Xu, W.; Zhang, C.; Liu, Q.; Liu, C.; Wang, S. Tree growth characteristics and flower bud differentiation of sweet cherry (Prunus avium) under different climate conditions in China. Hortic. Sci. 2010, 27, 6–13.
- Guler, Y.; Dikmen, F. Potential bee pollinators of sweet cherry in inclement weather conditions. J. Ento. Res. Soc. 2013, 15, 9–19. Available online: https://www.researchgate.net/publication/286268023 (accessed on 20 May 2022).
- Predieri, S.; Dris, R.; Sekse, L.; Rapparini, F. Influence of environmental factors and orchard management on yield and quality of sweet cherry. Food Agri. Envi. 2003, 1, 263–266. Available online: https://www.researchgate.net/publication/266226841 (accessed on 20 May 2022).
- Beppu, K.; Ikeda, T.; Kataoka, I. Effect of high temperature exposure time during flower bud formation on the occurrence of double pistils in ‘Satohnishiki’ sweet cherry. Sci. Hortic. 2001, 87, 77–84.
- Blanco, V.; Blayo-rose, P.J.; Torres-Sánchez, R.; Domingo, R. Influence of regulated deficit irrigation and environmental conditions on reproductive response of sweet cherry trees. Plants 2020, 9, 94.
- Zang, L.; Ferguson, L.; Whiting, M.D. Temperature effect on pistil viability and fruit set in sweet cherry. Sci. Hortic. 2018, 241, 8–17.
- Juhasz, A.; Sepsi, P.; Nagy, Z.; Tokei, L.; Hrotko, K. Water consumption of sweet cherry trees estimated by sap flow measurement. Sci. Hortic. 2013, 164, 41–49.
- Ranney, T.G.; Bassuk, N.L.; Whitlow, T.H. Osmotic adjustment and solute constituents in leaves and roots of water-stressed cherry (Prunus) Trees. J. Am. Soc. Hortic. Sci. 1991, 116, 684–688.
- Blanco, V.; Martínez-Hernández, G.B.; Artés-Hernández, F.; Blaya-Ros, P.J.; Torres-Sánchez, R.; Domingo, R. Water relations and quality changes throughout fruit development and shelf life of sweet cherry grown under regulated deficit irrigation. Agric. Water Manag. 2019, 217, 243–254.
- Ruiz-Sánchez, M.C.; Domingo, R.; Castel, J.R. Review. Deficit irrigation in fruit trees and vines in Spain. Span. J. Agric. Res. 2010, 8, 5.
- Vosnjak, M.; Mrzlic, D.; Hudina, M.; Usenik, V. The effect of water supply on sweet cherry phytochemicals in buds, leaf and fruit. Plants 2021, 10, 1131.
- Romano, G.S.; Cittadini, E.D.; Pugh, B.; Schouten, R. Sweet cherry quality in the horticultural production chain. Stewart Post Harv. Rev. 2006, 6, 2.
- Kazantzis, K.; Chatzicharissis, I.; Papachatzis, A.; Sotiropoulos, T.; Kalorizou, H.; Koutinas, N. Evaluation of sweet cherry cultivars introduced in Greece. J. Univ. Kraiova Ser. Hortic. 2011, 16, 293–296. Available online: http://pomologyinstitute.gr (accessed on 28 March 2022).
- Cherry growers Australia Inc. 2021, Varieties and Management, Cherry Quality. Available online: http://cherrygrowers.org.au (accessed on 21 May 2022).
- Clayton, M.; Biasi, W.V.; Agar, I.T.; Southwick, S.M.; Mitcham, E.I. Sensory quality of Bing sweet cherries following preharvest treatment with hydrogen cyanamide, calcium ammonium nitrate or gibberellic acid. HortScience 2006, 41, 745–748.
- Zang, C.; Whitting, M. Plant growth regulators improve sweet cherry fruit quality without reducing endocarp growth. Sci. Hortic. 2011, 150, 73–79.
- Kaiser, C.; Long, L.; Brewer, L. Understanding and Preventing Sweet Cherry Cracking. Oregan State University EM 9227. 2019. Available online: https://www.researchgate.net/publication/331358391 (accessed on 15 February 2022).
- Looney, N.E.; Lidster, P.D. Some growth regulator effects on fruit quality, mesocarp composition, and susceptibility to postharvest surface marking of sweet cherries. J. Amer. Soc. Hortic. Sci. 1980, 105, 130–134.
- Kappel, F.; MacDonald, I.K. Gibberellic acid increases fruit firmness, fruit size, and delays maturity of ‘sweetheart’ sweet cherry. Isr. J. Plant Sci. 2002, 56, 219–222.
- Einhorn, T.C.; Wang, Y.; Turner, J. Sweet cherry fruit firmness and postharvest quality of late maturing cultivars are improved with low rate and single application of Gibberellic Acid. HortScience 2013, 48, 1010–1017.
- Christensen, J.V. Rain-induced cracking of sweet cherries: Its causes and prevention. In Cherries: Crop Physiology, Production and Uses; Webster, A.D., Looney, N.E., Eds.; CAB International: Wallingford, UK, 1996; pp. 297–330. ISBN 9780851989365.
- Christensen, J.V. Cracking in cherries. III. Determination of cracking susceptibility. Acta Agric. Scand. 1972, 22, 128–136.
- Measham, P.F.; Bound, S.A.; Gracie, A.J.; Wilson, S.J. Incidence and type of cracking in sweet cherry (Prunus avium L.) are affected by genotype and season. Crop Pasture Sci. 2009, 60, 1002–1008.
- Winkler, A.; Knoche, M. Calcium and the physiology of sweet cherries: A review. Sci. Hortic. 2019, 245, 107–115.
- Winkler, A.; Fiedler, B.; Knoche, M. Calcium physiology of sweet cherry fruits. Trees 2020, 34, 1157–1167.
- Amarante, C.V.T.; Miqueloto, A.; Stefens, C.A.; Dos Santos, A.; Argenta, L.C. Changes in xylem functionality during apple fruit development: Implications on calcium concentration and incidence of bitter pit. Acta Hortic. 2013, 1012, 135–140.
- Xiloyannis, C.; Celano, G.; Montanaro, G.; Dichio, B.; Sebastiani, L.; Minnocci, A. Water relations, calcium and potassium concentration in fruits and leaves during annual growth in mature kiwifruit plants. Acta Hortic. 2001, 564, 129–134.
- Windt, C.W.; Gerkema, E.; Van As, H. Most water in the tomato truss is imported through the Xylem, not the Phloem: A nuclear magnetic resonance flow imaging study. Plant Phsiol. 2009, 151, 830–842.
- Winkler, A.; Peschel, S.; Kohrs, K.; Knoche, M. Rain cracking in sweet cherry is not due to excess water but localized skin phenomena. J. Am. Soc. Hortic. Sci. 2016, 141, 653–660.
- Blanco, V.; Zoffoli, J.P.; Ayala, M. Influence of high tunnel microclimate on fruit quality and calcium concentration in Santina sweet cherries in a Maditerranean climate. Agronomy 2021, 11, 1186.