1. TTFC Properties and Uses: CNS Delivery and Immunogenicity
In view of the numerous potential therapeutic applications of TTFC, especially in the fields of vaccination and neurology, different methods of production, characterization and evaluation were developed. This article therefore offers an overview of TTFC by describing its numerous properties regarding immunology and neuroprotection, but also reflecting the different methods used for producing it.
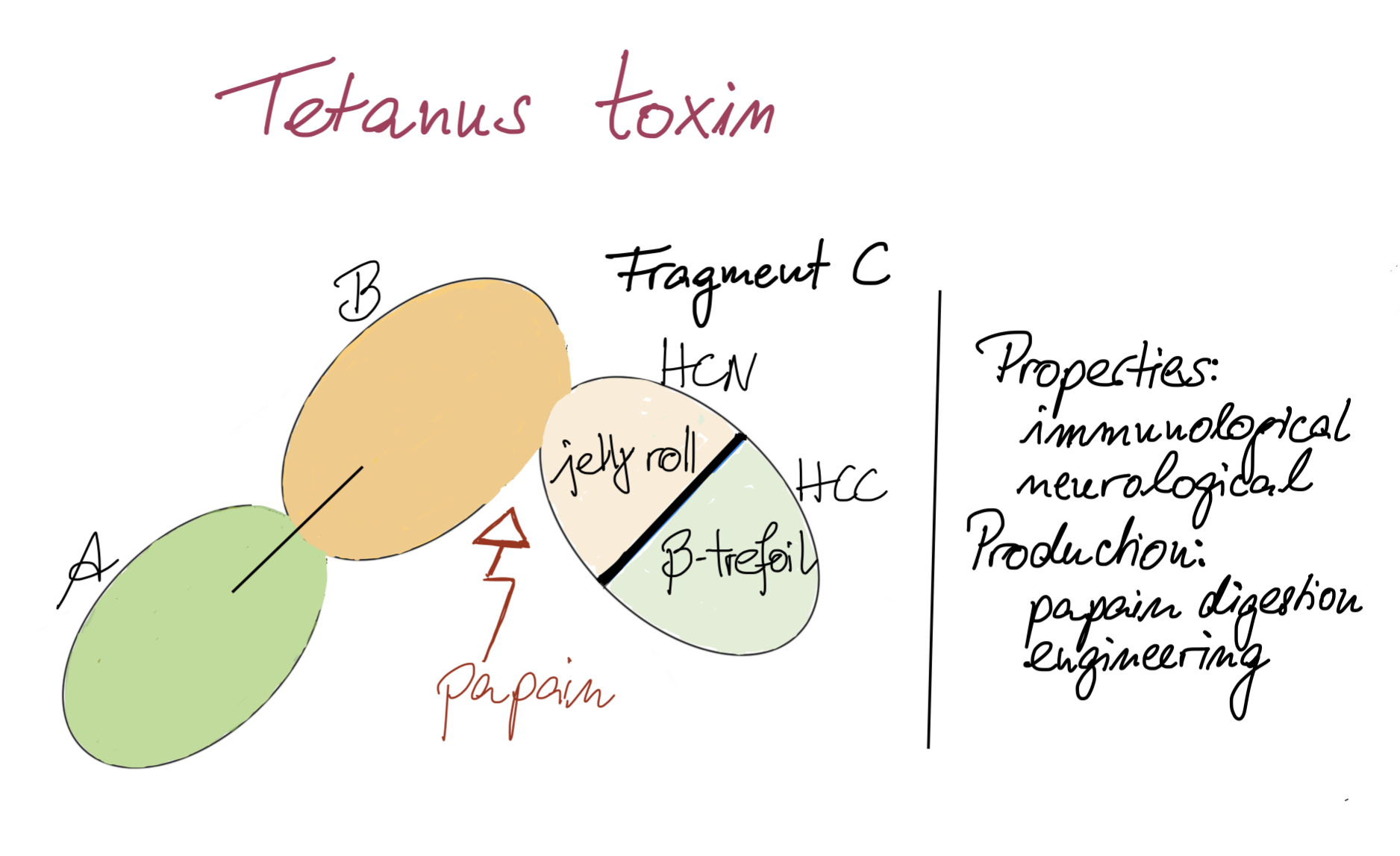
1.1. TTFC Neurological Properties
1.1.1. Permeability and CNS Delivery
TTFC was reported to bind to motor neurons (MN) [
19]. TTFC contains distinct binding sites, which are recognized by neuronal receptors. This is the first step of its transport into the CNS. It especially binds to polysialylgangliosides G
D1b and G
T1b, which are present in the CNS. Their role is to regulate protein activities at the lipid membrane [
20]. After ganglioside recognition, TTFC is internalized by MN at the NMJ [
21]. As it has the ability to form pores in lipid vesicles, the third step of its transfer is its translocation through the lipid membrane. This translocation depends on the ganglioside content but also on the pH of the membrane [
11,
22]. It finally reaches the neuronal perikaryon in the CNS, where it can target the recognition and the catalytic cleavage of neuronal substrates [
11]. Owing to its ability to reach the CNS and to specifically affect neurons, TTFC seems to be a promising candidate as a vector for neuronal disease treatment [
22].
Indeed, used as a fusion protein or carrier after chemical linkage, TTFC may be useful to deliver either proteins or drugs into the CNS to cure neurodegenerative diseases. After intramuscular injection or systemic administration, such chemically altered TTFC were shown to enhance protein delivery in neurons by 1000-fold in a mouse model [
23]. TTFC was also reported to improve both the neuronal uptake and the distribution of brain-directed therapeutics [
23,
24]. As the blood–brain barrier (BBB) is relatively difficult to cross, drugs are directly injected into the brain or into cerebrospinal fluid. Using TTFC as a vector for protein delivery could simplify the route of injection. Indeed, a fusion protein administrated by intramuscular injection was detected on endosomal and synaptic vesicles [
24]. These two locations are interesting as the synaptic membrane can be targeted for neurotransmitter-related protein or trophic factor delivery, and endosomal and lysosomal locations can be targeted for metabolic enzyme delivery [
23]. This method has been used successfully to deliver proteins to MN [
24]. Importantly, several studies have shown that a TTFC/cytokine fusion had no effect on neuronal properties or cytokine protection properties [
25].
Fusion proteins with TTFC can also be used to study synaptic connections. Fusion with a labeled protein (e.g., GFP, green fluorescence protein) allows transportation of the protein through synapses in a non-toxic way. Such a tool can be used to study the mechanisms of TTFC internalization and to map the neuronal network [
26]. Indeed, TTFC is able to fuse with other molecules without altering its trans-synaptic and retrograde properties. This type of fusion has led to the successful study of intra- and inter-neuronal trafficking in vivo [
27].
The unique properties of TTFC have also been exploited to construct a TT-derived peptide (Tet1-peptide, sequence HLNILSTLWKYR) as a new approach to delivering therapeutics to the CNS, as illustrated by the efficient delivery of small molecules into the CNS after intramuscular injection. This synthetic analog could thus also be used as a vector [
28].
1.1.2. Intrinsic Neuronal Protection
In addition to its carrier properties, TTFC has also been reported to display neuroprotective activity. Indeed, retrograde and trans-synaptic transport of TTFC into the CNS after muscular injection is similar to pathways followed by trophic factors [
29]. Different studies have demonstrated that TTFC had a trophic action in the brain and that it mimicked growth factors involved in survival pathways [
30]. Furthermore, TTFC prevents cell death by apoptosis via several routes. Briefly, apoptosis occurs after cytochrome c release and caspase-3 activation following induction by the apoptotic promoter Bax. This promoter, in turn, is activated upon reversion of Bcl-Xl activity (an anti-apoptotic protein) through the dephosphorylation of the
Bad gene [
31]. Caspases are involved in major apoptotic events such as DNA digestion, chromatin condensation or membrane blebbing. TTFC disturbs these events by inhibiting caspase-3 activity, thus preventing apoptotic cell death [
32]. TTFC also protects cells from apoptotic death by inhibiting Bad dissociation and association with Bcl-Xl. TTFC was thus shown to exert anti-apoptotic effects by inhibiting cell death pathways activated by Bad and by blocking pro-caspase-3 activation [
33].
Moreover, TTFC is also involved in the activation of survival signaling pathways. Trk receptors transmit their signals from axons to neuronal cell bodies in a retrograde manner. Neurotrophins binding to these receptors allow protein kinase activation and tyrosine residue auto-phosphorylation. These phosphorylated tyrosine residues are then recognized by intracellular signaling proteins activating a kinase cascade. For example, Trk activation can be the trigger for phosphoinositide 3-kinase (PI3K) pathway activation, which in turn activates protein kinase B (also known as Akt), which is necessary and sufficient for eukaryotic cell survival [
34]. TTFC stimulates signaling pathways Akt/PI3K, ERK and Raf/MEK/ERK [
30,
32,
34], which play a role in cell survival. For example, PI3K is involved in the prevention of low potassium-induced apoptotic death. TTFC can provide cell protection through its action on survival signaling pathways. TTFC also protects cells by activating p21Ras protein, which is essential for the inhibition of cell death pathways [
31,
32]. In addition, TTFC treatment affects calcium-related gene expression, suggesting that TTFC can impact anti-apoptotic pathways through calcium-related mechanisms [
29].
Another type of protection provided by TTFC concerns oxidative stress. Oxidative damage is observed in neurodegenerative diseases such as Parkinson’s disease (PD). Generation of reactive oxygen species can also contribute to MPP(+)-induced oxidative stress that activates a series of cellular factors that initiate apoptotic cell death [
31]. Cubí et al. showed that after the administration of TTFC, the ceramide content and the nSMase activity increased in cerebellar granule neurons and NGF-differentiated PC12 cells [
35]. It is well-known that an increase in ceramide level is related to neuron survival. Hence, as TTFC treatment increases ceramide contents, this provides neurons with a better protection against oxidation. TTFC is thus able to protect cells against neuronal oxidative stress and prevent oxidative damage.
Owing to its neuroprotective properties, TTFC has been widely tested in animal models of PD, cerebral ischemia, amyotrophic lateral sclerosis (ALS) or animals treated with molecules inducing apoptosis. TTFC was particularly reported to induce protection against dopamine (DA) loss and improved MN behavior [
30]. DA plays a key role in controlling locomotion and is the main neurotransmitter affected in PD. DA receptors are involved in the activation of Trk receptors, and it seems that the trkB pathway may be crucial for DA neuron survival. As mentioned above, TTFC has a neuroprotective effect, and that may be due to its known contribution to the activation of Trk receptors, thus improving mechanisms of DA neurotransmission [
33]. TTFC had also been identified as a neuroprotective agent in ALS animal models as it prevents anti-apoptotic effects [
30]. TTFC fusion proteins were tested in mice with ALS disease and displayed a combination of TTFC neuroprotective and carrier properties. Improvements in motor function activity were observed in TTFC fusion-treated mice, curbing disease progression and increasing the number of surviving MN [
29]. In parallel to MN survival, a reduction in microglial reactivity was detected, stopping disease progression. Another example worth mentioning was the use of naked DNA encoding TTFC (nDNA-TTFC) in the treatment of cerebral ischemia. This disease is characterized by a decrease in blood supply to the brain. This dysfunction can lead to permanent disability and death. There is currently no treatment to counter these serious effects. The efficacy of the treatment partly relies on the ability of the therapeutic treatment to reach the neurons after crossing the BBB. Every oxidative stress parameter was reduced after nDNA-TTFC treatment in all brains tested, protecting them from oxidative damage [
36]. This treatment may thus represent a non-invasive and non-viral therapeutic approach to treating cerebral ischemia. The unique properties of TTFC as a carrier and neuroprotector may pave the way for many applications to cure neurodegenerative diseases.
1.1.3. Overview of the Uses of TTFC for Its Neurological Properties
Based on all its properties, TTFC has already been used under different forms (e.g., alone, in fusion protein, conjugated) for neuronal applications, including in vivo in several studies, confirming its neurological properties. Examples of these applications are presented in Table 2 (see article).
1.2. Immunological Properties
1.2.1. Immunological Properties against Tetanus
Having presented the neurological properties of TTFC, we will now review its immunological properties. TTFC contains four universal epitopes of the 11 present in the TT sequence: 52 to 68 [
52], 83 to 103 [
52,
53,
54,
55,
56], 290 to 309/310 to 325 [
57] (these two peptide sequences may represent the same epitope) and 409 to 420 [
52] in the TTFC sequence. An immuno-informatics analysis performed by Nezafat et al. [
58] unveiled four regions of the TTFC as helper epitopes (53–69, 84–108, 220–247, 361–386).
According to other studies, TTFC also contains 13 of the 28 epitopes recognized by more than 75% of patients [
52,
53,
54,
55,
56,
57,
59]. This means that nearly half of TT epitopes are contained in the TTFC sequence. Ghafari et al. [
60] suggested that TTFC may be the immunodominant part of the toxin, i.e., that the fragment may elicit the humoral immune response. Moreover, TT neutralizing antibodies were shown to be mainly directed against TTFC in mouse and human models (100% for mouse models and 75% in a human model) [
61]. It is important to note that the many neutralizing antibodies described since the 1980s [
62,
63,
64] cannot individually neutralize TT in vivo. The development of an active and safe monoclonal antibody (mAb) remains a prospect for upcoming years [
65]. Hence, TTFC has become a very interesting candidate from an immunological point of view, as it could protect against tetanus. Ghotloo et al. [
66] gathered the state of knowledge on epitope mapping of TT and, more particularly, the epitopes localized on the TTFC by mAbs.
Keeping its immunological properties in mind, TTFC was then tested in vivo against tetanus. The first study conducted by Fairweather et al. tested the last 121 amino acids of the B fragment associated with TTFC in mice. One microgram of the fragment was injected without adjuvant and provided complete protection against TT, as all mice survived the TT challenge [
67]. Other studies were conducted on TTFC alone showing that it could induce a 13-month protection against TT when expressed in bacteria, yeasts, plant cells or insect cells [
68].
Further studies then compared its immunogenicity to the antigen present in the current vaccine (tetanus toxoid, TTd) [
69,
70]. Depending on the injection mode, the antibody response changed. No difference between TTd and TTFC was observed after the first transcutaneous delivery (injection of either 30 µg of each protein or molar equivalent doses). However, after the third injection, TTFC induced a 2 to 12 times higher antibody response (mainly IgG1 subclasses responses, IgA were not detected). Kinetics of antibody response of both proteins also seemed to differ. Using TTFC, a constant increase in protective response was observed, whereas using TTd, the level of protective response remained constant event after new injections. Using subcutaneous injection, contradictory results were obtained. In one study, a 100 times higher dose of TTd was required to induce the same protection as TTFC [
69], and in another, TTFC was 100 times less potent than TTd [
70]. The origin of these differences has not been fully investigated, but the induced protection seems to be linked to the mode of injection, which must be considered when developing new tetanus vaccines. Other modes of administration confirmed the capacity of TTFC to offer protection against tetanus: sublingual immunization using
Bacillus subtilis as an antigen delivery system showed full protection in mice [
71]. The capacity of TTFC to be used as an antigen in a tetanus vaccine has been demonstrated, but further investigations on the impact of the mode of injection must be conducted.
Because TTFC contains many epitopes, its use as a carrier to deliver molecules to the CNS may be delicate in vaccinated individuals. Studies have been conducted to analyze the immunogenic side effects of the administration of TTFC in vaccinated animals [
72]. They revealed that the amount of injected TTFC as a carrier (molar equivalent) generally exceeds 10,000 times the lethal dose of TT in a mouse model, and neutralization of all the TTFC injected was unlikely even for a vaccinated animal. This allowed a small part of TTFC to be internalized and to reach the MN. Results have shown that vaccinated and unvaccinated animals presented detectable retrograde transport of TTFC using fluorimetry [
72]. This demonstrated that at a high dose, TTFC could be used as a carrier for delivery of bioactive molecules into CNS even in vaccinated individuals. Another explanation could be linked to the mode of injection, as after an intramuscular injection, TTFC was not blocked by antibodies developed after immunization against TT. This could be due to the rapid uptake of the protein after intramuscular injection [
21].
Finally, as an attractive alternative to commercial ELISA assays, dipstick tests were developed using gold-conjugated TTFC [
73]. This conjugate was used to bind anti-tetanus antibodies in whole blood and plasma samples, with no cold chain requirement. The results obtained were highly promising, with a specificity greater than 0.9. The use of this device could thus be integrated into monitoring programs for populations at risk (non-immune or low titer individuals).
1.2.2. TTFC as a Fusion Protein: Enhancement of Immunogenicity
TTFC did not only induce protection against tetanus, it also enhanced immunogenicity of its partner when fused. DNA-based vaccines have shown their efficacy against infectious diseases but revealed disappointing results when used in tumor models. To overcome this problem, fusion with antigen helpers was developed. TTFC displays functional characteristics of helper antigens and was thus tested in several DNA fusion proteins [
74]. In particular, it improved T-cell immunity of viral oncogenes HPV-16 E6 and E7 (viral oncogenes targeted by tumor suppressor proteins p53 and pRb) used in DNA vaccines to protect against papillomavirus [
75]. The fusion with TTFC enhanced CD8+ T cell response against the protein of interest. Indeed, the mounting of a robust E7-specific T-cell response was observed after the administration of a TTFC fusion protein [
76]. This induced tumor regressing and prolonged survival of all mice tested. E7 stability was also improved by the fusion protein, as E7 accumulation was higher in cells treated with the fusion. This could be explained as TTFC contains universal epitopes for human CD4+ T lymphocytes: the CD4+ T-cell helper response produced after epitope recognition may promote CD8+ T-cell responses. In addition, TTFC has a stimulatory effect on IFN-γ and CD69 production, both involved in modulating the immune response. Enhancement of the immunogenicity using TTFC fusion proteins may thus be due to the action of these proteins [
60].
The fragment was also shown to enhance bacterial polysaccharide antigenicity in conjugate vaccines. As for TTd, TTFC has been tested as a carrier protein in conjugate vaccines, albeit because the production of TTFC does not require a formaldehyde detoxification step, its conjugation sites are easier to characterize [
77]. Several authors chose to use this protein to synthetize well-defined glycoconjugates to highlight the link between the conjugate structure and the induced immune response [
78,
79,
80,
81]. TTFC was shown to play a normal protein carrier role as its conjugation with the bacterial polysaccharides induced a higher and long-lasting immune response.
1.2.3. In Silico Design of Epitope-Based Vaccines
In the last decade, in silico methods have been used to design vaccines based on TTFC epitopes associated with other epitopes. For example, in the case of atherosclerosis, a multi-epitope construct was designed by fusing epitopes from TTFC with other immunogenic molecules such as calreticulin, heat shock protein 60, and cholera toxin B [
82]. Bioinformatics analysis allowed the authors to build a stable chimeric protein with the potential to shift the immune response and to reduce atherosclerosis. In silico approaches were also used to design anti-infective vaccines against brucellosis [
83] and leptospirosis [
84] in order to induce a strong immune response mediated by T- and B-cells. Immuno-informatics was also helpful to design a vaccine against melanoma [
85], by fusing several antigens, including TTFC. This multi-epitope vaccine (MEV) approach showed high immunogenicity, providing hope for cancer immunotherapy. In another study [
58], different algorithms and servers were used to design a novel MEV against cancer. The selected cytolytic T lymphocyte (CTL) epitopes were linked together to enhance epitope presentation. In parallel, different helper epitopes (including TTFC) were conjugated to stimulate helper T lymphocyte (HTL) immunity. All these epitopes were associated with heparin-binding hemagglutinin used as an adjuvant. The final protein was thus able to stimulate both cellular and humoral immune responses. An in vivo study was conducted to produce the corresponding vaccine in
E. coli in order to evaluate antitumor efficacy against the HPV-16 E7-expressing murine tumor cell line TC-1. The results obtained revealed a significantly higher IgG secretion with MEV containing TTFC epitopes compared to the E7 protein vaccine [
86].
1.2.4. TTFC Uses for Its Immunological Properties
As for its neurological properties, the immunological properties of TTFC have inspired many groups of research. Again, TTFC was used under different forms (e.g., alone, in fusion proteins, in conjugate vaccines) for diverse vaccine applications. Examples of these applications are presented in Table 3 (see article).
2. TTFC Production
2.1. Papain Digestion
One of the simplest ways of obtaining TTFC is to enzymatically digest TT. This can be achieved using the protease papain present in papaya fruits. Papain cleaves TT between Ser864 and Lys865 residues resulting in two fragments: TTFC and fragment A-B (
Figure 1). Papain was first used for this purpose by Helting et al. in 1974 [
7]. At that time, structural and functional studies of TT were difficult to ascertain because of the lability of the whole protein after purification. This is why researchers decided to split the protein into smaller fragments, trying to obtain separate structural information. The digestion was usually performed in 0.1 M phosphate buffer containing 1 mM EDTA and cysteine hydrochloride at pH 6.5. The experimental conditions (e.g., reaction temperature, amount of material) were progressively optimized [
7,
100,
101,
102,
103]. For example, in 2014, Murzello et al. [
100] digested TT within 30 min at 55 °C using 10 units of papain from papaya latex (Sigma P4762) per 10 mg of TT. To stop the reaction, the enzyme was inactivated by adding L-1-chloro-3-tosylamido-7-amino-2-heptanone and by letting the batch cool to 25 °C.
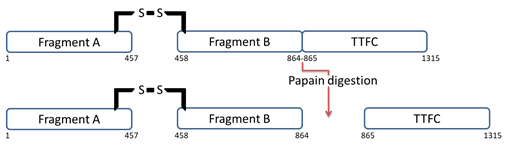 Figure 1.
Figure 1. TT structure before and after papain digestion. Papain digests the protein by splitting it into two fragments: TTFC and fragment A-B.
In the next step, TTFC was purified from the solution obtained after papain digestion. Usually, several chromatographic separations (e.g., size-exclusion chromatography) were performed to finally yield pure TTFC. Subsequently, an affinity chromatography column using an anti-fragment [A-B] antibody could complete the purification protocol. Sometimes, TTFC needed to be rechromatographed (on the same column material under different elution conditions) to remove the remaining contaminants. Yields obtained after all purification steps varied between 15 to 28% from the TT starting material (approximately 0.13 to 10 mg of protein per mL) [
7,
100,
101,
102,
103].
2.2. TTFC Production in Recombinant Systems
TTFC production using papain digestion was the first to be described, but as it was hazardous and a major difficulty was to eliminate the very small amount of undigested toxin from the fragment preparation, recombinant production systems were developed [
104]. These systems present some striking advantages compared to papain digestion. First, large amounts of TTFC can be efficiently produced since large numbers of host organisms (e.g., bacteria, yeast, plant cells, insect cells) can be grown in small- and large-scale cultures with standard laboratory equipment. Second, only controlled non-toxic proteins can be produced, which means that production is safer and toxic contaminations are easy to avoid [
105].
2.2.1. Escherichia coli as a Host for TTFC Production
In 1986, the cloning of the DNA encoding the TTFC from
C. tetani CN3911 strain was published [
106]. From this first study, several groups started to work on the production of TTFC as a heterologous protein in
E. coli (
Table 4, see article) [
104,
105,
106,
107,
108,
109,
110,
111,
112,
113]. Low amounts (from 1 to 10 mg/L) of recombinant proteins were obtained from expression vectors containing
trp or
tac promoters [
104,
105,
106]. TTFC was also produced as a fusion protein in the pMalc2x vector [
109]. TFC codon analysis revealed that the gene contained a high proportion of rare codons in
E. coli, and teams thus produced synthetic TTFC DNA in order to reduce rare codons and decrease the percentage of AT pairs, increasing protein production [
110,
111]. However, it appeared that the yield increased further when BL21 bacteria and an expression plasmid the including T7 promotor were used [
107,
108,
112,
113]. In 2011, Yu et al. [
110] optimized the fermentation production of 40 L to reach 333 mg/mL recombinant TTFC. As described recently [
114], the fermentation conditions were further optimized to increase the production of recombinant proteins. In the late 1990s, other bacterial expression systems were studied to produce and ensure the delivery of TTFC. In fact, these alternative bacterial expression systems were developed to produce oral vaccines against TT, i.e., they served both as an expression method and as a delivery system.
2.2.2. TTFC Expression and Delivery in Other Bacterial Host Strains
The production of heterologous proteins in cyanobacteria, phototrophic microorganisms with low nutrient requirements, may be an alternative. For eukaryote proteins, this is possible in the form of fusion proteins. Recently, in 2021, the TTFC protein was expressed in the cyanobacterium
Synechocystic sp. PCC6803 (
Synechocystis) [
115]. This bacterium was initially modified to stably express the tobacco etch virus protease (TEVp). Then in the
cpc operon locus, the native
cpcB gene encoding the abundant β-subunit of the phycocyanin was replaced with a fusion construct comprising the
cpcB and TTFC DNA sequences separated by a TEV cleavage site. In vivo cleavage led to the accumulation in the cytosol of soluble TTFC proteins and putative aggregates containing the uncleaved protein cpcB-tev-TTFC, the sum of the two proteins representing approximately 20% of the total protein content (TPC). Hidalgo Martinez et al. [
116] showed that the accumulation of soluble cpcB-tev-TTFC fusion proteins could occur as they contribute to the formation of phycobilisomes, comprising the major light-harvesting antenna complex for photosynthesis.
TTFC was produced by other bacterial recombinant expression systems for different applications. All these bacteria were not only used to express TTFC but also to deliver it in oral vaccination approaches. These delivery systems were developed in order to produce safe, stable and inexpensive oral vaccines. Such vaccines are particularly appropriate in developing countries because syringes and needles used for injected vaccination may be used repeatedly and lead to the transmission of infectious agents such as HIV, hepatitis B and C viruses. Oral immunization also simplifies the administration of vaccines, allowing less qualified health workers to immunize populations. These vaccines are highly compatible with mass immunization programs as their logistics are simpler. Finally, oral administration is usually preferred by both children and adults compared to parenteral injection [
117]. The objective of oral vaccines is to pass through the intestinal mucosal surfaces to reach the immune system. Mucosal surfaces play a role in nutrient uptake, and an immune response against these dietary antigens is unwanted and generally suppressed to avoid food intolerance. It is thus difficult to induce a systemic immune response after the delivery of antigens to these surfaces. The immune response can, however, be improved by associating the antigen with a bacterium [
118]. Lactic acid bacteria are well adapted to deliver antigens through mucosal surfaces and are interesting since they are generally recognized as safe (GRAS) by the FDA [
119]. These nonpathogenic food-grade Gram-positive bacteria can be resistant to the harsh conditions of the intestinal environment. For example, the
Lactococcus lactis bacterium can prevent direct contact of antigens with gastric acid and proteolytic enzymes, conferring a higher resistance in the intestinal medium [
120]. Different lactic acid bacteria were tested as expression systems for TTFC:
L. plantarum,
L. lactis and
L. casei [
118,
119,
120,
121,
122,
123,
124,
125,
126,
127]. Generally, the recombinant plasmids were constructed in the
E. coli DH5α host strain and were subsequently introduced into alternative bacterial host strains by electroporation. Several strains and plasmids were tested to express the protein, leading in most cases to successful intragastric immunization. Strains UCP1060 and MG1363 were most often used for
L. lactis, strains NCIMB8826 for
L. plantarum and strains ATCC393 for
L. casei [
118,
119,
120,
121,
122,
123]. Plasmids pMEC46 and pMEC127 were both frequently used within these strains, and the administration of MG1363(pMEC46) and NCIMB8826(pMEC127) strains, in particular, showed a high antibody production in mice [
119,
121,
123]. To increase the immune response, different mutations were also tested in bacterial strains. For example,
L. plantarum and
L. lactis (Alr
−) mutant strains (replacing of L-alanine by D-alanine), both producing TTFC intracellularly, were more immunogenic than their wild-type counterparts [
121]. These expression systems were good alternatives to attenuate pathogenic bacterial delivery systems such as
Salmonella typhimurium [
128] and
Mycobacterium bovis BCG [
129].
In summary, several bacterial production systems have been developed to produce TTFC. Different modifications have been made to the plasmids and the host strains to improve the fragment expression. Bacterial expression systems are widely used because they are cheap, their genetic manipulation is easy, and these fermentation systems are often easy to scale up. However, expressing foreign antigens in bacterial strains also has its drawbacks: Gram-negative bacteria cell walls contain lipopolysaccharides (LPS), which are toxic pyrogens. Consequently, proteins expressed in these bacteria must be carefully tested on LPS content before being used. Yeasts or plant cells exhibit no such features and could thus be attractive hosts for TTFC expression.
2.2.3. Recombinant TTFC Expression in Yeast and Plant Cells
The yeast species
Saccharomyces cerevisiae and
Pichia pastoris were used for recombinant TTFC expression as well as tobacco chloroplasts, representing a common plant expression system. To obtain satisfying levels of expression in yeast, a synthetic TTFC gene was constructed, containing a codon-optimized high CG gene lacking the fortuitous polyadenylation sites, which gave rise to truncated mRNA [
142,
143]. Around 60 to 90 mg/L of soluble protein were obtained using
S. cerevisiae S150-2B strain expressing the synthetic TTFC gene (pWYG5-TET15) [
142]. However, in
S. cerevisiae, the recombinant TTFC secreted in the culture medium was immunogenically inactive due to glycosylation. A methylotrophic yeast,
P. pastoris, produced a much higher yield of 12 g/L of soluble protein using the integrative plasmid with the promotor from the methanol-induced alcohol oxidase gene and the synthetic TTFC gene (pPIC3-TET15) [
143]. When the native gene was cloned into a yeast expression plasmid (p9k-G) with the alpha factor secretion signal and expressed in
P. pastoris, the secreted TTFC was glycosylated. Site-directed mutation performed on five potential N-glycosylation sites showed that the number and localization of these sites impact the expression and secretion of TTFC. Indeed, decreasing the number of N-glycosylation sites decreased TTFC secretion [
144].
Conversely, plant cells did not require the use of the synthetic TTFC gene to obtain high yields of proteins. Indeed, the AT-rich bacterial gene, when expressed in a cassette consisting of a PrrnLT7g10 cassette (plasmids pJST10 and pJST11, respectively), produced twice as much protein (25% of total cell protein (TCP)) than the high-GC synthetic gene. However, this high level of TTFC is detrimental to plants, as evidenced by the chlorotic phenotype in plants [
145,
146]. The chloroplast expression system has inherited similarities with the prokaryotic expression systems derived from the ancestral cyanobacterium. They contain a plastid-encoded RNA polymerase, which can be compared to the bacterial RNA polymerase [
147]. Two types of methods can be used to produce proteins in plant expression systems: either nuclear transgenic plants (low expression levels generally achieved) or DNA can be introduced into the chloroplast genome by particle bombardment (higher expression levels of recombinant proteins) [
148]. Proteins are usually extracted from leaf tissues using an extraction buffer. In addition, chloroplasts are heat stable, cheap to produce, and they can remove undesirable selectable markers and use operons for multi-antigen expression; they produce a high level of immunogenic recombinant proteins, specifically target the gene and retain the gene product in the plastid; this system also benefits from environmental containment as a result of maternal inheritance [
148]. Michoux et al. studied the production capacity of the
Nt-pJST12 line obtained from a plasmid expressing the synthetic gene in a cassette consisting of a PrrnLatpB cassette. They showed that 92 g of TTFC could be produced annually, using five 250 L-bioreactors and running 20 rounds of production per reactor per year. This quantity of TTFC should be enough to launch preclinical and clinical trials [
149]. Chloroplast-expressing systems were able to yield high amounts of protein but were not always efficient as vaccines. As both plant and the expressed vaccine are degraded in the intestinal tract, oral immunization was inefficient [
150]. Nevertheless, intranasal immunization was protective in mice against the TT challenge [
145,
146].
Beyond yeast and chloroplasts, insect cells were also tested as host organisms using baculovirus expression vector p36C to co-infect
Spodoptera frugipedra cells. The resulting TTFC was soluble and showed an immunogenic activity, but its secondary structure was different from the TTFC obtained in
E. coli as its ganglioside binding properties were altered [
151].
TTFC can be produced using a panel of different production methods. Purification [
104,
105,
107,
108,
110,
152] and characterization [
108,
110,
122,
133,
152] steps are then usually the last ones performed in protein production to finally obtain the required quality. In 2020, Chai et al. established a simple method to purify TTFC by ion-exchange chromatography [
153].
The development of new vaccines and neuroprotective agents present major issues in drug discovery. New vaccines must be well defined and better controlled, present great antigenic and immunogenic activities and be as stable as possible to allow mass vaccination campaigns in developing countries. The development of neuroprotective agents mostly focuses on the drug delivery into neuronal cells as the BBB is difficult to cross. TTFC protein can be a good candidate to overcome problematics in both fields. Its structure has been characterized allowing a better understanding of its biological properties. Recombinant TTFC is easily characterized and was shown to be sufficient for the protection of mice against tetanus. Its properties can also be used to protect against other diseases (e.g., multiple myeloma, cholera) as TTFC enhances immunogenicity of therapeutic proteins when fused to them. This fragment is responsible for neuronal binding of TT, meaning that it can reach neurons. This property has been widely studied for drug delivery into the CNS, as this is a key point in the cure of neurodegenerative diseases (e.g., ALS, AD, PD). These studies have shown that when conjugated or fused to a drug, TTFC retains its neuronal binding capacity and is a good carrier for drug delivery into the brain. Intrinsic neuroprotective properties were also discovered confirming the growing interest for this protein in the neurological field. TTFC thus appears to be a promising protein for drug discovery; as its production, characterization, and properties are well described. Further studies must now be conducted on different animal models to confirm its biological relevance and hopefully will result in the development of new drugs. In addition, TTFC-based immune-informatics approaches also seem to be an attractive and effective design to access new therapeutic entities against certain zoonotic diseases (e.g., brucellosis, leptospirosis).
In conclusion, TTFC protein can be considered to be a useful drug discovery toolbox. Few proteins offer such a wide range of uses including as a carrier, delivery vector, conjugate, booster, inducer, and neuroprotector. With the development of nanomedicines, and consideration for brain safety [
154], the use of TTFC could pave the way for novel, safer applications.
This entry is adapted from the peer-reviewed paper 10.3390/ph15060756