1. Introduction
Human beings are higher-order organisms living with the microbial world inside and outside of their bodies. Microbes have inhabited the earth for more than a million years before humans [
1]. The genome of microorganisms and humans collectively constitutes the human microbiome [
2]. The human body is a habitat for various microorganisms such as bacteria, viruses, fungi, and parasites. Almost every place in the human body acts as a distinct microbial niche. The main sites of microbial colonization are the airways, urogenital tract, eyes, skin, oral, pulmonary tract, and gastrointestinal (GI) tract [
3,
4]. In particular, the human GI tract fosters millions of bacteria, a greater number than that fostered by the eukaryotic cells of the human body [
5].
1.1. Gut Microbiota
The gut microbiota (GM) has been categorized into autochthonous microbes and allochthonous microbes, with different functions. The former type resides in the epithelial layer of colonic mucosa, whereas the latter one passes through the lumen [
6]. The prevalence of GM is not alike everywhere in the GI tract; the distribution varies across the intestinal tract, with preference for three microhabitats: floating cells in the intestinal lumen, adherent cells in the mucosal layer, and intestinal epithelial cells [
7]. GM varies from the duodenum to the ileum, with high microbial loads in the colon region [
8]. Although there is a great diversity of microbes residing in the human gut, most of the microbiota are from phyla Firmicutes and Bacteroidetes [
9]. About 1% of microbes are from other phyla, including Proteobacteria, Actinobacteria, Fusobacteria, Verrucomicrobia [
10], Bifidobacteria, Lentisphaerae, and Spirochaetes [
11]. The availability of microbes in the GI tract depends on changes in pH, nutrient availability, GI transit, mucin secretion, immune functions, and the host’s age and health [
10,
12].
The GI tract executes various functions such as digestion of food, absorption of nutrients and water, protection against pathogens, and energy balance, etc. The GM and their metabolites modulate GI functions, such as providing mucosal immune function, intestinal permeability [
13], sensitivity [
14], enteric nervous system (ENS) activity [
15], visceral pain [
16]; brain functions and behaviors, including emotions [
17], pain [
18], and stress responses [
19]; and in brain biochemistry [
20]. Neural networks control the physiological functions of the GI tract. The autonomic nerves connect the central nervous system (CNS) with the gut [
21]. The gut is innervated by the ENS, which is connected to the CNS and allows the exchange of metabolites and information [
22].
1.2. Gut Microbiota, Neuronal System, and Neurodegenerative Diseases (NDs)
The CNS modulates the GI tract and the ENS through the network of vagus nerves, with sympathetic and parasympathetic nerves of the autonomic nervous system (ANS), hypothalamic–pituitary–adrenal (HPA) axis, gut hormones, and cytokines [
23]. The CNS modulates the GM through the enteric environment and signaling molecules [
24]. The GM performs various functions such as nutrient metabolism, synthesis of vitamins, breaking down of drugs, promoting intestinal barrier [
25] and maintaining the production of short-chain fatty acids (SCFA). During these functions, the GM releases small molecules and metabolites that can stimulate mucus secretion [
26]. In addition to these functions, the GM triggers the innate immune system and leads to the development of gut-associated lymphoid tissue (GALT), which results in the development of adaptive local immunity [
27].
The function of the nervous system is disturbed if there are any physiological interruptions in the GM, which can be termed gut dysbiosis. Gut dysbiosis can directly modify the function of the immune system and tissue barriers, such as the blood–brain barrier (BBB) [
28], which in turn influences the brain functions, and results in stress response, cognitive activities, and changes to behavior and memory [
18,
20], possibly leading to depression [
29] and anxiety [
30]. The GI functions depend on the interactions of the gut microbes with the brain, through the creation of a “microbiota–gut–brain axis” network [
28]. Studies showed that GM takes part in the physiology of the brain. In a case where there are any interruptions in the microbiota by any harmful nutrients, microbes can elicit different signaling pathways, including oxidative stress, energy metabolism, mitochondrial function, and neuroinflammation, and disturb the epigenetic mechanisms, which eventually influence gene expression [
31].
The gut and brain connections are mediated by various microbial metabolites called neuromodulators [
28,
31]. In addition to maintaining immune and metabolic health [
4], the gut–brain axis performs various other functions such as brain development [
32] neurogenesis [
33], and the CNS–ENS interactions in NDs and neuroinflammation diseases such as Parkinson’s disease (PD), Alzheimer’s disease (AD), multiple sclerosis (MS), autism spectrum disorder (ASD) [
34], and major depressive disorders (MDDs) [
27].
1.3. Parkinson’s Disease (PD)
Among other NDs, patients with PD showed GI disturbance even before developing motor symptoms [
35]. About 80% of PD patients are observed with clinical GI symptoms such as nausea, vomiting [
36], and constipation [
37]. Along with these symptoms, stomach and colon motility disturbances are very common in PD [
38]. The GM communicates with the brain and vice versa. Any changes in the gut–brain axis may interfere with the function of each other [
39].
The clinical manifestations of PD and the GM–brain axis need to be well characterized, including GI modulations and immunological and neuroendocrine mechanisms. Many extra-neuronal factors, such as nutrition and environmental factors, may cause impacts on metabolism, the immune system, distress to the brain physiology, and neuronal function that leads to neuropathogenesis [
31]. Nutritional habits greatly affect GM’s colonization, maturation, and changes throughout human life [
40]. The ecology of GM changes according to age, diet, medications, and geographical location [
41].
2. Pathophysiology of Parkinson’s Disease (PD)
The etiology of PD is multifaceted, including genetics, aging, gut dysfunction, and environment [
124]. Familial PD can occur because of the point mutations in the alpha-synuclein (α-syn) gene, and locus duplication, triplication and sporadic PD may be due to genetic and environmental factors. Pesticides such as rotenone, paraquat, and 1-methyl-4-fenyl 1,2,3,6-tetrahydropyridine are one of the causative agents of PD [
124].
James Parkinson first described PD in his “An Essay on the Shaking Palsy” in 1817. He originally described the muscular weakness and non-tremulous form of PD [
125]. Many decades later, after Charlot’s study in 1957, the cause of PD was recognized as a loss of neuronal cells in substantia nigra [
126]. In 1960, the neurotransmitter dopamine was found to be diminished in the striatum of PD patients [
127]. The other contributing factors for the fall of dopaminergic and non-dopaminergic neurons in the brains of PD patients include misfolded proteins, ubiquitin-proteosome, and autophagy lysosomal system errors, increased oxidative stress, mitochondrial dysfunction, and inflammation [
128,
129].
PD affects several brain regions, including pigmented nuclei in the midbrain, brainstem, olfactory tubercle, cerebral cortex, and some of the peripheral nervous system [
130]. The degeneration of dopaminergic neurons of the substantia nigra compacta (SNc) region and their projections to the striatum developed during the disease progression; due to this degeneration of neurons of limbic portions of the striatum, the motor signs of PD are visible before the non-motor signs [
131]. In addition to the loss of dopaminergic neurons, serotonergic cells in the median raphe, noradrenergic cells in the locus coeruleus, and cholinergic cells of the nucleus basalis are also involved to a lesser extent.
Most PD is idiopathic, dominantly inherited genetic variants associated with intraneuronal Lewy bodies inclusions. Some other genetic causes for PD, such as autosomal-dominant and recessive susceptibility genes, have now been identified. Mutations in the α-syn gene were the first identified genetic cause of PD. Other most common mutations are in the gene glucocerebrosidase (GBA) and the lysine-rich repeat kinase 2 (LRRK-2) gene [
132]. α-syn is involved in the mitochondrial function and synaptic plasticity, highly concentrated in the nerve terminals and abnormally aggregated as Lewy bodies, which are the prominent component of PD [
132]. Impaired proteasomal degradation of ubiquitin-C-terminal hydrolase-L1 and β-glucocerebrosidase [
133,
134] results in intraneuronal accumulation, misfolding, and phosphorylation at serine-129 of α-syn [
135]. The toxic aggregation of α-syn affects the nigrostriatal dopaminergic neurons and induces dopamine autotoxicity [
136].
The studies showed that the α-syn aggregation could be initiated in the gut and olfactory bulb [
137]; within the GI tract, the deposited α-syn showed a rostro-caudal gradient and was highly concentrated in the submandibular gland and lower in the esophagus [
138]. Later, the vagus nerve spreads the α-syn to the brain stem and toward the cortex [
137]. Thus, α-synucleinopathy causes neurodegeneration, impaired axonal transport, and degradation of synaptic terminals [
139]. A recent study showed the involvement of microRNAs (miRNAs) in PD pathogenesis [
140]. Parkin, DJ-1, PINK1, ATP13A2, DNAJC6, PLA2G6, SYNJ1, FBOX7, SNCA, LRRK2, and VPS35 are considered risk factors for PD [
141,
142,
143,
144,
145,
146,
147]. Endogenous toxins such as misfolded or aggregated proteins, synuclein and
tau [
148,
149], and pro-inflammatory cytokines secreted by T lymphocytes and glial cells, are also associated with PD pathogenesis [
150,
151].
The four important clinical features of parkinsonism are TRAP: tremor at rest, rigidity, akinesia (bradykinesia), and postural instability [
152]. Bradykinesia is an easily recognizable symptom of PD, observed with decreased neuronal density in the substantia nigra, which results in disturbed motor activities [
153,
154]. Tremor is one of the most common PD symptoms and involves involuntary movement of hands, lips, chin, jaw, and legs [
155]. Clinical-pathological studies revealed that the neurons in the subgroup of the midbrain degenerated in PD patients with tremors [
156]. The next cardinal symptom is rigidity, characterized by resistance and pain [
157,
158]. PD patients may also show postural deformities with flexed neck, trunk, elbows, and knees [
159,
160]. Some skeletal deformities such as neck flexion, truncal flexion, scoliosis [
161,
162], and extreme flexion of the thoracolumbar spine (camptocormia) are also common in PD [
162]. The late stage of PD is characterized by postural instability, which may cause hip fractures [
163]. Freezing, which is akinesia, means the complete loss of movement [
164], affects the legs, arms, and eyelids [
165], and is reported more frequently in men than in women [
166]. In addition, some of the non-motor symptoms such as autonomic dysfunction, cognitive disorders, and sleep abnormalities are common features of PD [
156].
The bidirectional communication between gut and brain in the case of PD is represented by an integrative organization of both intrinsic and extrinsic nervous systems [
167]. PD shows GI dysfunction and cardiovascular, urogenital, thermoregulatory, sleep, and respiratory abnormalities [
168]. The PD lesions initiated in the ENS later involve the CNS or vice versa. α-syn lesions are transmitted to the midbrain via the nasal olfactory lobe, and to the temporal lobe and GI system via the ENS [
169].
According to Braak’s hypothesis, pathogens may enter through the oral, nasal, or digestive tract, and reach the gut, initiating Lewy bodies in case of sporadic PD [
124]. Then, Lewy bodies occur in the dorsal motor nucleus (DMN) of the vagus in the medulla oblongata, vagus nerves, and anterior olfactory nucleus. The Lewy bodies spread within the CNS, substantia nigra, locus coeruleus, neocortex, mesocortex, and prefrontal cortex [
170] (
Figure 3).
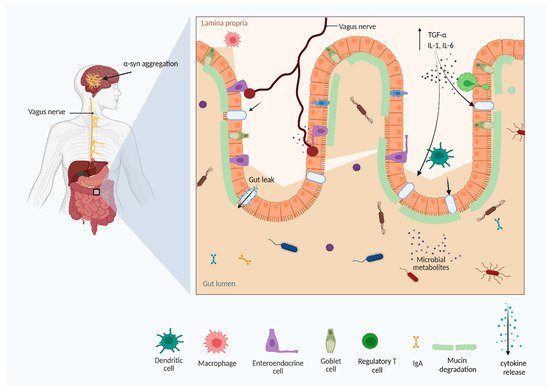
Figure 3. The gut dysbiosis and Parkinson’s disease. Gut dysbiosis and defects in intestinal barrier function facilitate the release of material metabolites, endotoxins, and other antigens into the gastrointestinal system, which further activates the immune system and the release of pro-inflammatory cytokines. Chronic immune activation may cause neuroinflammation and neurodegenerative diseases (Figure created using BioRender.com).
2.1. Toll-Like Receptors Expression in PD
Toll-like receptors (TLRs) belong to the pattern recognition receptors family [
171]. TLRs are associated with activating the immune cells and initiating the immune response. Different types of T and B lymphocytes were found to express a variety of TLRs [
172]. TLRs recognize antigens and cause immune responses through antigen-presenting cells. TLRs were also found to regulate the functions of CD4+ and CD25+ Treg cells (regulatory T cells) and modulate the immune response [
173]. In the case of gut dysbiosis, GM trigger inflammatory effects against Lewy bodies and its derived antigens via CD4
+ T-cell response, causing early gut inflammation, which progresses to PD [
174]. In PD, the hyperphosphorylated α-syn aggregates in the brain [
175] stimulate innate immune responses in the microglial cells [
176] by increasing the TLRs expression in the glial cells [
177], especially in the substantia nigra region [
178]. TLRs can act as lipopolysaccharides (LPS) receptors, a cell-wall component of gram-negative bacteria. It binds the LPS and activates the pro-inflammatory and anti-microbial cytokines [
179].
Higher TLRs expression was found in the brain of α-synucleinopathies such as PD, dementia with Lewy bodies, and multiple system atrophy, and is involved in the innate immunity, which becomes a therapeutic target for these disorders [
180]. TLRs’ activation causes neuroinflammation, stimulates NF-κB and pro-IL-1β, and triggers NDs [
181]. TLR2 and TLR4 are the target for PD, as both these receptors are involved in PD development [
182]. Aggregated α-syn can bind to the TLR2 and TLR4 and initiate the immune responses in PD. TLR2 can bind with the oligomeric α-syn, resulting in pro-inflammatory signals and neurodegeneration [
183]. TLR4 binds with any monomeric or oligomeric α-syn and helps in clearing the α-syn, thereby exerting an alternative role of neuroprotection [
184]. α-synucleinopathy studies in rat models evaluated that blocking TLR2 and activating TLR4 provide protective functions [
185,
186] (
Figure 4).
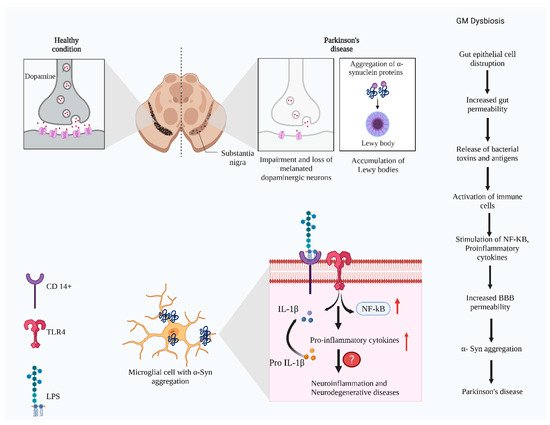
Figure 4. The representation of molecular signaling and progression of Parkinson’s disease in substantia nigra. Gut dysbiosis activates the immune system. The released cytokines may disturb the blood–brain barrier, facilitating the entry of bacterial metabolites and other antigens to the central nervous system. It causes α-syn aggregation in the substantia nigra of the brain. Hyperphosphorylated α-syn recruits the TLR4 and CD14+ in the microglial cells, promoting the neuronal immune responses by activating the NF-κb, pro-IL β pro-inflammatory cytokines. The exact mechanism and the players are not elucidated completely (Figure created using BioRender.com).
3. Signaling Pathways Associated with Gut Microbial Changes and PD
The GM and the CNS are linked through neural, endocrine, and immune signaling. For example, the microbiota in the gut can induce the cells to synthesize neurotransmitters and digestive hormones, which could alter the brain and behavior [
187]. In turn, the CNS can control the GM through adrenergic nerve signaling by regulating the neurotransmitters on immune mediators that shape the GM [
187]. Microbiota could affect the hypothalamus-pituitary-adrenal (HPA) axis [
188]. Another important metabolic mediator of neuroimmune function are the SCFAs. SCFAs regulate the CNS by modulating microglia [
189]. SCFAs signal through the G protein-coupled receptors (GPR), free fatty acid receptors (FFAR), and GPR109. The GPR109 receptor binds to butyrate and induces the production of IL-10-secreting Treg cells [
190]. PD patients reported a low abundance of
Prevotella and SCFA-producing
F. prausnitzii and
Clostridium IV, and lowered SCFA metabolites in their gut [
191,
192,
193,
194]. A study in α-syn-overexpressing mice demonstrated that the microbial products could be involved in the pathogenesis of PD, by regulating the immune cells in the brain [
195]. GM play a prominent role in amino acid metabolism, influencing neuroinflammatory diseases [
196]. The intestinal microbiota mediate the endocrine, immune, and neural pathways. Accordingly, gut dysbiosis can alter behavior, mood, and neuroinflammatory responses through the HPA axis, lipopolysaccharide, neurotransmitter, and SCFAs [
196] (
Figure 5).
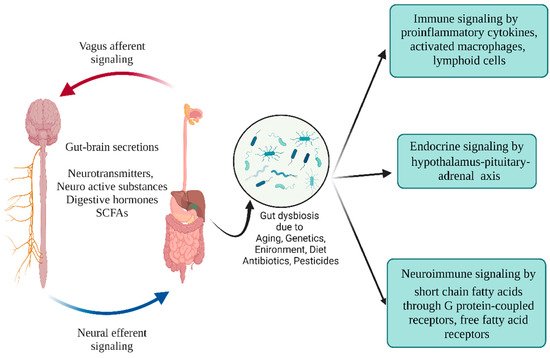
Figure 5. Schematic illustration of endocrine, immune, and neuroimmune signaling pathways. Gut microbes help maintain intestinal integrity by balancing the microbial products, neurotransmitters, and SCFAs across the enteric and immune systems. Microbial dysbiosis triggers activated immune cells, macrophages, dendritic cells, and pro-inflammatory cytokines. (Figure created using BioRender.com).
4. Gut Microbiota and Parkinson’s Disease
GI dysfunction has been recognized as associated with PD pathogenesis [
197]. GM and its metabolites interfere with the host’s behavior, immunity, cognition, and metabolism [
198,
199,
200,
201]. The changes in the GM composition and its metabolites have been identified as a vital reason for the induction and progression of PD [
202]. The bidirectional communication between the gut–brain axis is possible through GM [
203]. The afferent fibers from the gut are connected to the anterior/posterior, cingulate, cerebral, amygdalar, and insular cortices, and the efferent fibers project to the gut’s smooth muscles [
203]. The studies showed that the GM of PD patients were rich in Enterobacteriaceae, which was related to the severity of postural instability and gait difficulties (PIGD) in PD patients, which indicates a positive correlation with PIGD [
191].
In the meantime, the abundance of Lactobacillaceae and the decreased abundance of Prevotellaceae in the gut are related to reducing the intestinal hormone ghrelin, a regulating component of nigrostriatal dopamine (DA) [
204]. GM regulated the synthesis of DA by controlling the DA-producing enzymes [
205]. The majority of DA was produced by the GM, for example,
Bacillus spp. [
18].
Members of Prevotellaceae are reduced in PD patients, which might reduce the mucin synthesis in the gut mucosal layer. The reduction of mucin leads to increased gut permeability, simplifying the entry of bacterial toxins and antigens, thus favoring the aggregation of α-syn in the colon and brain [
169,
206]. Another possibility of α-syn accumulation is decreased butyrate synthesizing bacteria and increased pro-inflammatory Proteobacteria, which trigger inflammation-induced α-syn misfolding [
192].
Clostridium IV,
Clostridium XVIII,
Holdemania,
Aquabacterium,
Sphingomonas,
Butyricicoccus, and
Anerotruncus were found in the feces of PD patients and are negatively associated with the disease duration [
207]. A reduction in the abundance of Prevotellaceae, Lachnospiraceae, Lactobacillaceae, and Streptococcaceae was reported in PD patients [
208].
PD patients are reported to have enteric problems such as bacterial growth in the small intestine, especially
Helicobacter pylori infection, and constipation.
H. pylori infection increases the risk of PD and worsens motor symptoms [
209].
Faecalibacterium spp. level was reduced,
Ralstonia spp. were significantly increased, and there was no change in the
Bifidobacterium in the mucus of PD patients [
210].
The abundance of Enterobacteriaceae,
Lactobacillus,
Escherichia,
Shigella,
Streptococcus,
Proteus, and
Enterococcus were increased, while
Clostridium coccoides,
Bacteroides fragilis, Bacteroidetes, and Prevotellaceae were decreased in Chinese PD patients. In addition, the cellulose degraders
Blautia,
Faecalibacterium, and
Ruminococcus were significantly decreased in PD patients compared to healthy controls [
193,
194,
211]. GM dysbiosis reduces mucin production and increases intestinal permeability, related to PD progression and development [
151].
Another study in a mouse model of PD revealed that the hydrogen sulfide produced by
Prevotella protects the dopaminergic neurons [
212]. Dysbiosis of gut microbes reduces the SCFAs, which possibly induces changes in the GI motility and the ENS [
193], and increases the neurotoxin and endotoxin production, which can eventually lead to PD development [
211]. GM metabolism is responsible for the changes in β-glucuronate and tryptophan degradation pathways in PD [
213].
A high abundance of Akkermansia, Bifidobacterium, Ruminococcaceae, and Lactobacillus were found in the fecal microbiome of PD patients, which increases the xenobiotic degrading pathways in PD [
214]. Cellulose-degrading bacterial genera such as
Blautia,
Ruminococcus, and
Faecalibacterium, and pathogens
Streptococcus,
Escherichia,
Shigella,
Enterococcus, and
Proteus were increased in PD subjects compared to normal subjects [
211]. In contrast, Qian et al. reported that
Bacteroides plebeus (
B. plebeus),
B. coprocola,
B. dorei,
B. massiliensis,
P. copri,
Dorea longecatena,
Faecalibacterium,
Stoquefchus massiliensis,
Coprococcus eutactus, and
Ruminococcus callidus abundances were decreased. Members such as
Christensenella minuta,
Christensenella hongkongensis,
Catabacter,
Lactobacillus mucosae,
Oscillospira,
Bifidobacterium,
Ruminococcus bromii, and
Papillibacter cinnamivorans richness were increased in Chinese PD patients [
215]. The members of Christensenellaceae, Verrucomicrobiaceae, Lactobacillaceae, Bifidobacteriaceae, Lachnospiraceae, and Pasteurellaceae were abundantly found in Russian PD patients [
216]. The increase in abundances of
A. muciniphila,
P. copri, and
Eubacterium biforme was observed in American PD patients [
217]. The bacterial genera responsible for anti-inflammatory and neuroprotective effects (
Butyrivibrio, Pseudobutyvibrio,
Coprococcus, and
Blautia) were reduced in Italian PD patients [
215]. Relative abundances of anti-inflammatory butyrate-producing bacteria (
Blautia,
Coprococcus,
Roseburia, and
Faecalibacterium) were decreased, and the pro-inflammatory bacteria (
Ralstonia) were found to increase in PD patients [
192]. GI complications, dysbiosis, immune dysregulation, and inflammation were found in PD and inflammatory bowel diseases [
218,
219]. PD patients are often diagnosed with enteric problems such as constipation, and are reported to have less abundance of
Bifidobacterium,
Prevotella, and
Lactobacillus, and are rich in Firmicutes [
220].
Aging is a physiological function [
221] that induces an imbalance in the pro-inflammatory and anti-inflammatory changes and has been a major factor in various human diseases [
222]. Aging reduces the abundance of
Bifidobacteria,
Lactobacilli, and SCFA-producing
Faecalibacterium prausnitzii,
Eubacterium spp.,
Roseburia spp., and
Ruminococcus spp. [
223]. Aging-induced microbiota changes before the gut dysfunction could disturb various signaling pathways through the microbial metabolites and can negatively affect neurodegeneration [
224]. The accumulation of aging-related somatic damages and impaired cellular repair mechanisms can develop PD. The compensatory cellular repair mechanisms, such as mitochondrial oxygenation, ubiquitination, proteolysis, and autophagy processes were reduced during aging, resulting in increased radical production, oxidative stress, genomic instability, and DNA mutations [
225], leading to abnormal deposition of brain proteins [
226]. Aging can cause axial impairment of gait and postural control in PD [
227].
This entry is adapted from the peer-reviewed paper 10.3390/microorganisms10081544