Your browser does not fully support modern features. Please upgrade for a smoother experience.
Please note this is an old version of this entry, which may differ significantly from the current revision.
The acronym MSC refers either to: Mesenchymal Stem Cell, a term popularized by Caplan in the 1990s and broadly used in the past decades, or Multipotent Mesenchymal Stromal Cell, which is the terminology promoted by Mesenchymal and Tissue Stem Cell Committee of the International Society of Cell Therapy
- immunity
- mesenchymal stem cells
- stromal cells
1. Definition of MSC
The acronym MSC refers either to: Mesenchymal Stem Cell, a term popularized by Caplan in the 1990s [1] and broadly used in the past decades, or Multipotent Mesenchymal Stromal Cell, which is the terminology promoted by Mesenchymal and Tissue Stem Cell Committee of the International Society of Cell Therapy [31].
The adjective “mesenchymal” (“middle” in Greek) suggests a putative mesodermal origin of MSCs, implying that MSCs may derive from the “middle” layer during the embryonic development [1]. A neuroectodermal NC origin of MSC has been highlighted in many tissues by several investigators [4,12,15,16,17,32,33,34]. NC cells have the capacity to migrate and to participate in the organization of ectodermal and endodermal tissues [33]. This migration ability may be retained during adulthood, by homing to injured tissues [1,6,20,35,36]. Even if MSCs terminology and definition have been a matter of debate (notably about their “stemness”), there is a consensus on certain elements.
First, MSCs of the BM are distinguishable from hematopoietic stem cells, because of their ability to adhere to plastic vessel and to grow [1,31,37,38,39]. Morphologically, MSCs are spindle-shaped cells that after several passages bear a more homogeneously fibroblastic phenotype [40].
Second, MSCs are characterized by a singular expression of surface proteins. Among the classical markers of MSCs, we aforementioned CD90 (or Thy-1), CD105 (or endoglin, TGF beta receptor), and CD73 (or ecto-5′-nucleotidase) [8,37,38,41]. Other markers have been described for MSC such as CD140b (or PDGFR beta), CD271 (or low-affinity NGFR) [8,42,43], CD146 (MUC18), and CD248 (or endosialin/tumor endothelium marker 1) [7,9,22]. However, these markers are not specific to MSCs and can be found on other cell types. Additionally, MSCs are negative for CD45 (pan-leukocyte marker) and CD34 [37,38], even if some investigators evoked that CD34 may be expressed but lost in ex vivo culture expanded MSC [8,44]. Actually, it is important to note that the in vitro expression of some markers does not always correlate with their expression patterns in vivo [40].
Third, MSCs are multipotent progenitor cells, able to differentiate at least in vitro into three different subsets: adipocyte, chondrocyte and osteoblast [6,37,38]. In addition to these cell types, other possibilities of differentiation have been evoked [45]. Pericytes in the arteries can acquire a macrophage-like phenotype even with phagocytic properties [46,47]. This phenotype switching involves the transcription factor Krüppel-like factor 4 (KLF4) and has been associated with development of atherosclerosis. Some culture conditions can promote smooth muscle and striated muscle gene expression, whereas others promote cardiac or liver gene expression [48]. Finally, glial/neuronal differentiation potential has also been reported [40,41] and which may be due to the NC origin of specific subsets of MSC [7,11,12,15,16,17,19].
MSCs have been isolated from various tissues [4,5,6,40] and including: bone-marrow [49], adipose tissue [50,51], lungs [52], synovial membrane [53], kidney [6], liver [54], dental tissue [55,56,57,58,59], cord blood [60,61], and amniotic fluid notably [62]. We highlight that different names have been attributed to these cells depending on their tissue locations (Table 1).
Table 1. The different names given to Multipotent Mesenchymal Stem/Stromal Cells (MSC) and their tissue localization.
| Tissue | Human | Reference |
|---|---|---|
| Bone Marrow | Bone Marrow-Mesenchymal Stem Cell or Adventitial Reticular Cell or Myoid Cell |
[17,63,64,65] |
| Adipose tissue | Adipose Stem Cell | [50,51] |
| Intervertebral disc | Skeletal Progenitor Cell | [66] |
| Synovial membrane | Synovial Membrane-derived MSCs Fibroblast-like synoviocytes (FLS) |
[53,67] |
| Amniotic fluid | Amniotic Stem Cell | [62] |
| Cord blood Placenta |
Umbilical Cord Blood Stem Cell or Wharton’s Jelly derived MSC |
[60,61] |
| Dental tissue | Dental Pulp Stem Cell or Stem cells from Human Exfoliated Deciduous teeth or Periodontal Ligament MSC or Stem Cell from Apical Papilla or Dental Follicle Precursor Cell |
[55,56,57,58,59] |
| Kidney | Mesangial Cell | [4] |
| Brain | Pericytes, perivascular fibroblasts | [68,69] |
| Liver | Hepatic Stellate Cell (HSC) or Perisinusoidal Cell or Ito Cell |
[54,70] |
| Lung | Human Bronchial Fibroblasts or Lung-resident MSC |
[52] |
2. Origins of Pericytes/Perivascular MSC-Fibroblasts Derived from the Neural Crest and/or Mesoderm Embryonic Tissues
The ontogeny of MSC before they rich their final position in adult tissues is still a matter of debate [4]. The identification of MSC relies on the characterization of genetic and protein markers (e.g., tyrosine kinase PDGF α or β receptors, Schwann cell myelin P zero, glioma-associated transcription factor Gli1, collagen, Acta2/alpha SMA, Cspg4/Neuronglial 2-NG2, CD146/Melanoma cell adhesion molecule, CD248/Tumor Endothelial marker 1-TEM1) not restricted to MSC but also shared with pericytes (first identified by the French scientist Charles Rouget in 1873), vascular smooth muscle cells (VSMC), and perivascular fibroblasts [4,5,6,9,11,22,71]. Of note, the latter cell subset do not really fit the definition of pericyte because they are not embedded in the vascular basement membrane [72].
It is now generally accepted that a large pool of MSC is found essentially at the perivascular level and with morphology and marker expression profile similar to pericytes [5,73]. Several studies have shown that post-capillary venule pericytes from the bone marrow are able to differentiate into differentiated MSCs such as osteoblasts and chondrocytes in vivo [74]. More recent genetic lineage-tracing experiments and single-cell RNA sequencing data has reinforced a close link between pericytes and MSC phenotypes, particularly in the CNS, a tissue where the highest density of pericytes has been found in the body. RNA profiling of mouse brain vasculature revealed a rather unique pool of perivascular cells made of a two pericyte clusters and three subsets of perivascular fibroblasts [69,75]. Pathway and gene ontology enrichment analyses revealed that fibulin+ type I fibroblasts are the main subtype involved in ECM production and fibrosis. The type III (cell migration-inducing protein-CEMIP+ perivascular fibroblasts) showed robust expression of various growth factors, including VEGF-A. Interestingly, the type I to type II (potassium calcium-activated channel subfamily M alpha 1-KCNMA1+ fibroblasts) trajectory was continuous with pericyte type 2 suggesting a lineage from type I to type II to pericytes and consistent with a study in zebrafish demonstrating the stem cell potential of perivascular fibroblasts to differentiate into pericytes [76]. It was estimated that ten perivascular fibroblasts were present per intersegmented vessel but only less than 10% of these cells could differentiate into pericytes. Garcia et al. further discussed the possibility that type II perivascular fibroblasts in the brain probably represent an intermediate state exhibiting a transitional mural cell transcriptional phenotype [69].
MSC have several different developmental origins as reviewed by Majeski [77]. The majority of the MSC/pericytes in the head region, including the CNS, are neural crest (NC) derived, as demonstrated in chick-quail chimeras carried out initially by the French scientist Nicole le Douarin and colleagues [78]. In the peripheral nerves, NC will give rise to perineurial fibroblasts and Schwann cells [79]. More recently, two independent studies published in 2017 have suggested that brain pericytes could also be derived from mesoderm-derived myeloid progenitor cells [80,81]. Studies on the thymus demonstrated that perivascular MSC/pericytes are derived from the NC [32,82,83]. The origins of pericytes in the gut [84], lung [85] and liver [13,86] have been mapped to the mesothelium although NC can give rise to MSC-like cells in the gut [87].
In the kidney, the metanephric mesenchyme of the intermediate mesoderm will give rise to nephrons (from the distal convoluted tubule to the podocytes) and also to all major stromal interstitial cells, including the pericytes, perivascular fibroblasts, VSMCs, and mesangial cells [88]. a NC origin of a subset of perivascular fibroblasts of the kidney and contributing to fibrosis has been proposed from genetic lineage tracing experiments using the Schwann cell Myelin P Zero promoter-GFP/LacZ mice [12]. In the aorta, MSC may have at least four different developmental origins, secondary heart field, NC, somites, and splanchnic mesoderm. This invasion of mesothelial cells occurs at about the same time as the appearance of primitive endothelial and hematopoietic progenitors within the splanchnopleura. The primitive endothelial cells (EC) within the splanchnopleura colonize the floor of the aorta and differentiate in situ to produce the vasculature of the body wall, kidney, visceral organs, and limbs [77]. This process of vasculogenesis involving PDGF high expression by EC is consistent with the notion that mesothelial-derived MSCs are localized to BM via the invasion of the vasculature. Coronary vessels in the heart appear to have a similar development [89]. Mesothelial cells are known to undergo epithelial-to-mesenchymal transition (EMT) to delaminate and to migrate into the organs to produce their mesenchymal components. Interestingly, recent studies also point to a close ontogenic relationship between pericytes/VSMC and perivascular fibroblasts in many organs and supporting the current paradigm of such relationships in pathological settings for instance in the brain and lungs. The recent studies preach for the existence of a continuum of pericytes/perivascular MSC-fibroblasts cell phenotypes observed along vessels (and possibly nerves) and which suggest that these cells can (trans)differentiate into each other in conjunction with vessel/axonal/tissue remodeling. However, this interesting and promising paradigm requires further investigation.
The close relationship of MSC and progenitors with the vasculature will endow them as a possible source of new cells for physiological turnover for the repair or regeneration of local lesions. The canonical and current scenario is that damage to any tissue would release the MSC from its perivascular niche, they will divide and secrete immunoregulatory and trophic factors. Different signaling mechanisms may govern MSC mobilization from the perivascular niche, detaching from the endothelial cues and invading the parenchyma in response to injuries. This is exemplified by the importance of PDGF-B/PDGFRβ which has been demonstrated in many organs such heart, lung, and gut [90].
To date, we have a better idea into the embryonic origin of pericytes/perivascular MSC-fibroblasts in different organs but it is still critical to decipher the mechanisms governing their proliferation spreading along (as well evading) growing vessels in conjunction with angiogenesis (Figure 1). The capacity of these cells to circulate in the blood in various disease settings is of great and emerging importance from a clinical standpoint and including the identification of novel predictive soluble biomarkers of an ongoing pathological process in the tissues. Indeed, CD45−CD31−PDPN+ proinflammatory mesenchymal, or PRIME, cells have been identified in the blood from patients with rheumatoid arthritis, and these cells shared features of inflammatory synovial fibroblasts and predicting inflammatory flares [91]. This line of future studies is of great importance in cancer and other chronic inflammatory diseases associated with infectious diseases. In the context of cancer, some studies relate to the capacity of NG2+ pericytes to give rise to mesenchymal tumors (i.e., osteosarcoma) [92].
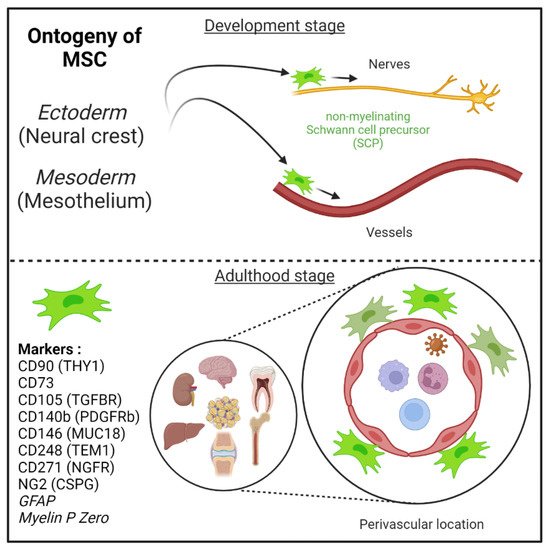
Figure 1. Mesenchymal stem cells (MSC) are derived from either he embryonic ectoderm (neural crest) or the mesoderm. MSCs can migrate along nerves and vessels during development and reside in virtually all post-natal organs and tissues. Along the nerves, MSCs are also known as non-myelinating precursor Schwann cells. Their location around vessels to form perivascular immune privileged niches has been demonstrated by several teams. MSC express several canonical markers which are differentially expressed in all major organs. CD271, GFAP, and MPZ are canonical neuroglial markers.
MSCs can migrate along nerves and vessels during development and reside in virtually all post-natal organs and tissues [6,15,16] (Figure 1). Along the nerves, MSCs are also known as non-myelinating precursor Schwann cells [19,93]. Their location around vessels to form perivascular immune privileged niches has been demonstrated by several teams [4,5,6,9,35,36,94,95,96]. Moreover, perivascular MSC may be able to sense and respond to an event (e.g., virus) in the local environment, via their ability to promote tissue immunoregulatory activities (see below) [36,44,97,98]. Perivascular MSC may therefore represent ideal tissue sanctuaries for viruses remaining dormant while protected from immune attack and rebounding particularly in the context of immunosuppressive drug treatments.
3. Role of MSC in Health (Immunoregulatory Activities and Tissue Repair) and Diseases (Fibrosis)
As described in the model of da Silva Meirelles et al. proposed in 2006, the MSCs in the vascular wall of tissue contribute to tissue maintenance (Figure 2) [6].
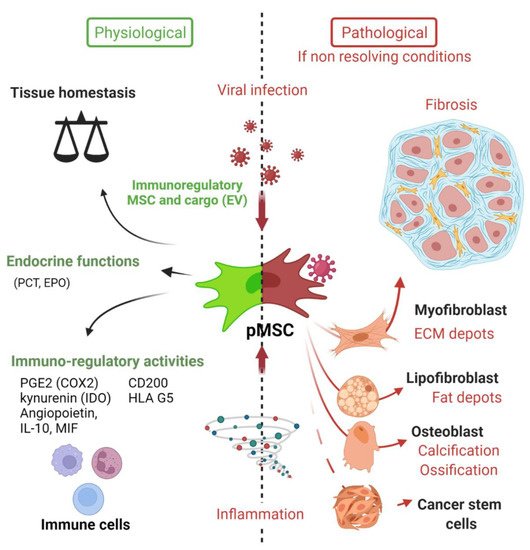
Figure 2. Deciphering the role of Mesenchymal stem cells in the context of viral infection. In physiological conditions, MSC have important immune functions to control viral infection as glatekeepers around vessels (perivascular MSC) and capable of mounting an innate immune antiviral response. MSC will also promote release of cytokines and chemokines to recruit immune cells to clear pathogens. Equally important is the expression of many immune regulatory factors to terminate the adaptive immune response to limit further cell injuries and promote tissue repair. Many viruses may infect directly MSC in tissues, thus remaining in an immunoprivileged niche favoring virus persistence, spreading and possible virus rebound in immunocompromised patients. Viruses associated to chronic inflammation (non-resolving) may also affect MSC differentiation (e.g., into myofibroblast) leading to excess of extracellular cell matrix production and contributing to organ dysfunction. Importantly, allogenic MSC and derived extracellular vesicles (EV) are nowadays important immunoregulatory cargo injected to patients for the treatment of inflammatory-infectious diseases such as COVID-19. Safety issues are nevertheless highly warranted. PCT (pro-calcitonin); EPO (erythropoietin) hormones.
Their motility ability allow them to migrate to the site of inflammation or injury to establish an appropriate response [1,6,20,35,36]. This response indubitably involves their immunoregulatory activities, that have already been comprehensively reviewed by several teams [10,36,44,98,99,100,101]. Among these well-known activities, MSCs modulate innate immune response by different mechanisms: decreasing Dendritic Cells (DCs)’ antigen presentation capacity, maturation and cytokine secretion [97,102]; reducing neutrophils burst respiration [103] and apoptosis; inhibiting natural killer (NK) cell proliferation, cytotoxicity and cytokine production [104,105,106,107]; inhibiting pro-inflammatory factor secretion by activated macrophages [97,108,109,110]. MSCs also exert immunoregulatory activities on adaptive immunity by: reducing T cell proliferation [44,111]; promoting T cell shift from pro-inflammatory (Th1) to anti-inflammatory (Th2) states [97], inhibiting cytotoxic CD8+ T cells [112,113]; inhibiting B cell proliferation through direct effect and T cell-mediated inhibition [114,115,116]; inducing the proliferation of regulatory T cells (Treg) [117,118,119,120].
These actions on the immune system involve both contact-dependent and contact-independent mechanisms. Indeed, MSCs have been shown to have effects on cell survival, function and proliferation of various immune cells by directly interacting with cell surface molecules and receptors [98]. For example, MSCs were shown to inhibit both T cell and B cell proliferation by the interaction between programmed cell death protein 1 (PD1), expressed by lymphocytes, and its ligand PDL1 expressed by MSCs [111]. Additionally, Fas (CD95)—Fas ligand (CD95L) axis is engaged and can induce inflammatory T cell apoptosis [121]. MSCs have also been shown to express the molecule CD200 (named OX2 in rodents) and regulating DC as well as macrophage/microglia immune cell activation via CD200R [122,123,124,125]. Liu and colleagues provided recently a very comprehensive review on the role of many more immunoregulatory cell surface ligands and receptors (e.g., Galectin1, 3, 9) allowing MSC to control directly innate and adaptive immune cells [124].
Contact-independent mechanisms involve the release of soluble immunoactive substances and extracellular vesicles (EVs), which form the MSC-derived secretome. Prostaglandin E2 (PGE2) and indoleamine 2,3-dioxygenase (IDO) (or nitric oxide, NO in mouse) expressed by MSC have been largely described for their immunosuppressive properties, for comprehensive review [10]. The chemokine—iNOS—IDO axis mediated by MSCs leads to T cell inhibition (Shi et al. [10]). Additionally, PGE2 production by MSCs leads to IL-10 secretion by M2-polarized macrophages [110], inhibition of DCs [97], reduction of NK cell activity [105], inhibition of Th17 cells and induction of Tregs [126]. Of note, other immunomodulatory factors have been reported to be secreted by MSCs: TGF-β1, hepatic growth factor (HGF), metalloproteinase-modified CCL2 (mpCCL2), leukemia inhibitory factor (LIF), and human leukocyte antigen-G5 (HLA-G5) [44].
Moreover, mounting evidence in recent years suggests an important role of EVs containing non-coding RNAs, including miRNAs, in the regulation of the immune system by MSCs [127,128].
The recruitment of immune cells to the injury site is granted by the release of soluble factors that leads to cell chemotaxis. Of note, MSCs can participate to this phenomenon by secreting a broad range of chemokines: CCL2, CCL3, CCL4, CCL5, CCL7, CCL20, CCL26, CXC3CL1, CXCL5, CXCL11, CXCL1, CXCL2, CXCL8, CCL10, and CXCL12 [10,44]. It is important to note that immunoregulatory activities of MSC will be markedly upregulated in response to the canonical IFN-gamma produced by T and NK cells [10].
Of further note, MSCs were initially thought to be important in regenerative medicine due to their ability to differentiate into multiple cell lineages, thereby supporting tissue repair. However, studies of past decades have found that regenerative activities on injured tissue were more likely associated with the MSC-derived secretome rather than the differentiation potential of engrafted MSCs [41,100,129,130]. Indeed, during a lesion, diverse mechanisms governed by MSCs-derived secretome participate in tissue regeneration and homeostasis [36,41,44]. Among them, we can cite: (1) anti-apoptosis effects mediated either by vascular endothelial growth factor (VEGF), HGF, insulin-like growth factor-1 (IGF-I), stanniocalcin-1, TGF-β, basic fibroblast growth factor (bFGF) or granulocyte-macrophage colony-stimulating factor (GM-CSF); (2) anti-scar with bFGF and HGF; (3) support and growth of tissue progenitor cells mediated by stem cell factor (SCF), LIF, SDF1 alpha/CXCL12, macrophage colony-stimulating factor (M-CSF) and angiopoietin-1; (4) angiogenesis stimulated by bFGF, VEGF, placental growth factor (PIGF), CCL2, interleukin 6 and ECM [44]. Finally, as aforementioned MSCs are able to mediate tissue regeneration by EVs release [131,132,133,134,135]. These EVs contain notably micro-RNAs that target pathways involved in angiogenesis and tissue remodeling [130].
4. MSCs’ Pathological Contributions (Fibrosis and Vessel Calcification)
Despite the beneficial activities that would turn MSCs into an obvious therapeutic approach [2], MSCs are also involved in pathological processes such as cancer [136,137,138] or fibrogenesis (Figure 2) [9,22,23]. In order to maintain the integrity of an organ, a fibrous scar composed of collagen is formed, leading (if uncontrolled) to chronic inflammation, fibrosis and to a loss of organ function [139]. Scar-forming cells are myofibroblasts, which were at length thought to be from an epithelial origin, after epithelial-mesenchymal transition (EMT) [24,140,141]. However, mounting evidence is pointing out the role of resident perivascular MSCs in myofibroblast differentiation and expansion, for comprehensive review, [9,14]. Perivascular MSC (also named pericytes by some authors) are nowadays more and more widely evoked as the main source of collagen-producing cells in fibrosis [11,94,142,143,144,145,146], notably thanks to genetic fate tracing experiment [12,14,143,147,148]. The role of MSCs in giving rise to myofibroblasts has been validated in several models of tissue fibrosis as reviewed by El Agha and colleagues in 2017. For instance, resident perivascular MSCs (genetically traced using hedgehog transcriptional activator glioma-associated oncogene homolog 1 (Gli1)+ or MPZ+ promoters to drive fluorescent protein expression, e.g., GFP) and its profibrotic activity have been described in murine model of fibrosis in either the kidney [12,147], lungs [147,149], heart [147,150], liver [147,148] or bone marrow [151].
Blood vessel remodeling can occur across a variety of pathologic conditions, including osteogenesis-like calcification (arteriosclerosis) and atheroma plaque formation (atherosclerosis) [183,184]. Although some reports have suggested that adventitial fibroblasts can contribute to pathologic changes within the vessel intima and media [185], it has now been shown, using GLI-1 line tracing experiments, that MSC can contribute to vessel calcification through a process of transdifferentiation into osteoblast-like cells [186].
The pathological role of MSCs is mostly associated with an initial and unresolved injury. Among the causes of injury, we will now focus on viral infection and how it affects resident MSCs in the different organs.
This entry is adapted from the peer-reviewed paper 10.3390/ijms23148038
This entry is offline, you can click here to edit this entry!