Fibrosis results from defective wound healing processes often seen after chronic injury and/or inflammation in a range of organs. Progressive fibrotic events may lead to permanent organ damage/failure. The hallmark of fibrosis is the excessive accumulation of extracellular matrix (ECM), mostly produced by pathological myofibroblasts and myofibroblast-like cells. The Hippo signaling pathway is an evolutionarily conserved kinase cascade, which has been described well for its crucial role in cell proliferation, apoptosis, cell fate decisions, and stem cell self-renewal during development, homeostasis, and tissue regeneration.
1. Introduction
Impaired tissue repair processes due to the dysregulation of molecular and cellular events after organ injury lead to organ fibrosis, a common cause of organ failure after chronic injury and/or inflammation [
1]. A hallmark of fibrosis is the excessive deposition of the collagen-rich extracellular matrix (ECM), especially due to an imbalance between collagen synthesis and remodeling. One of the key processes in fibrosis is the differentiation/transition of fibroblasts into myofibroblasts, the main fibrotic cellular phenotype, which produces a series of ECM components such as collagens, laminins, and fibronectins [
2,
3]. Numerous cell types, including epithelial and endothelial cells are also able to transdifferentiate into myofibroblastic phenotype (such as epithelial-mesenchymal transition and endothelial-mesenchymal transition) during organ fibrogenesis [
4]. Recent studies have implicated the contribution of Hippo signaling pathway components in the fibrosis of various tissues, including the lung [
5,
6,
7], liver [
8,
9,
10,
11,
12,
13,
14], kidney [
15,
16,
17], and heart [
18,
19,
20]. The Hippo signaling pathway was first identified and illustrated in
Drosophila. In mammals, the key components of the pathway are highly conserved, including several serine/threonine kinases and transcriptional factors, which are orthologs of
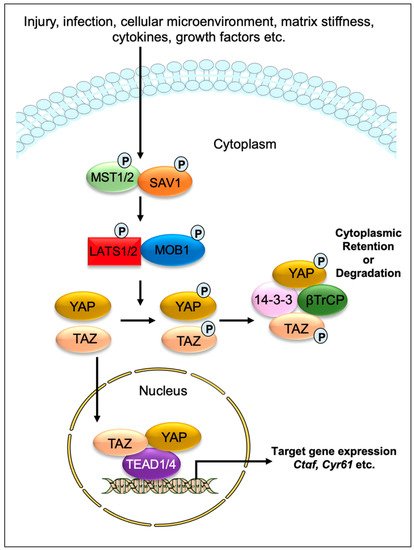
Drosophila proteins. The transcriptional regulator Yes-associated protein (YAP) and its co-activator PDZ-binding motif (TAZ/WWTR1) are the main effectors of this pathway. In response to physiological or pathological stimuli, sterile 20-like protein kinase (MST1/2) forms complexes with the adaptor protein Salvador 1 (SAV1) that phosphorylates large tumor suppressors (LATS1/2) and LATS1/2-interacting protein MOB kinase activator 1 (MOB1). The phosphorylated LATS1/2–MOB1 complexes, in turn, phosphorylate YAP and TAZ, resulting in the cytoplasmic retention or polyubiquitination and subsequent degradation of YAP/TAZ by proteasomes during autophagy (
Figure 1). In contrast, dephosphorylation of upstream kinase cascade drives the nuclear trafficking of YAP and TAZ, where they can interact with numerous transcription factors, including TEA domain DNA-binding family members (TEAD1–4), and regulate the expression of Hippo pathway target genes responsible for cell proliferation, survival, and differentiation [
21,
22,
23].
Figure 1. Graphical demonstration of the Hippo signaling pathway. Various physiological and pathological signals can induce the Hippo signaling pathway. In mammals, the core elements of the Hippo pathway mainly consist of serine/threonine kinases, transcriptional factors, and their cofactors. The transcriptional factors Yes-associated protein (YAP) and its coactivator PDZ-binding motif (TAZ/WWTR1) are the key effectors of the Hippo pathway. After physiological or pathological stimuli, sterile 20-like protein kinase (MST1/2) interacts, phosphorylates, and forms complexes with the adaptor protein Salvador 1 (SAV1), which phosphorylates large tumor suppressor (LATS1/2) and LATS1/2-interacting protein MOB kinase activator 1 (MOB1). The phosphorylated LATS1/2–MOB1 complex then phosphorylates YAP and TAZ, which promotes cytoplasmic retention or polyubiquitination and consequent degradation of YAP/TAZ by proteasomes during autophagy. However, dephosphorylation of upstream kinases leads to the nuclear translocation of YAP and TAZ, where they can interact with various transcription factors including TEA domain DNA-binding family members (TEAD1–4) and regulate the expression of Hippo pathway target genes such as Ctgf and Cyr61.
The Hippo signaling pathway has been extensively studied for its role as an organ size controller that regulates cell proliferation, apoptosis, cell fate decisions, and stem cell self-renewal during development, homeostasis, regeneration, and cancer formation in numerous mammalian organs [
21,
22,
24,
25,
26]. A growing number of studies have reported that the cascade components also play a critical role in fibrotic diseases. For example, the nuclear YAP/TAZ in resident cardiac fibroblasts obtained from preclinical myocardial infarction (MI) models [
18,
19,
27], are sufficient to direct the transdifferentiation of cardiac fibroblasts into pathological myofibroblasts [
27,
28]. Similarly, YAP/TAZ are important regulators of pathological fibroblast activation in pulmonary fibrosis [
5] and hepatic stellate cell activation in liver fibrosis [
11,
12,
13,
14]. Furthermore, activated YAP/TAZ in interstitial myofibroblasts promotes kidney fibrosis [
16], while hyperactivated YAP/TAZ in non-fibroblast cells, such as macrophages [
29,
30], epithelial cells [
31,
32,
33], and hepatocytes [
8,
10] contribute to the pathogenesis of fibrosis. Consistent with these findings showing the fibrotic effects of YAP/TAZ, their inactivation was found to be beneficial for preventing myofibroblast formation and fibrosis development [
34]. Like YAP/TAZ, Hippo kinases are also known to contribute to a range of fibrotic diseases [
20,
35,
36,
37,
38]. As such, treatment of mice with an MST1/2 blocker XMU-MP-1 inhibits pressure-induced cardiac hypertrophy and fibrosis (
Table 1) [
39]. Similarly, LATS1/2-mutant cardiac fibroblasts are less prone to develop into fibrotic phenotype after infarction-induced injury [
35], while deletion of LATS1/2 stimulates YAP/TAZ activation in myofibroblasts and aggravates kidney fibrosis [
40]. Similarly, genetic inactivation of SAV1 in renal tubule cells promotes renal interstitial fibrosis [
36]. In fibrotic diseases, various profibrotic signals can induce the function of the Hippo effectors. For example, stiff substrates drive the nuclear localization of YAP and TAZ in fibroblasts and regulates their differentiation into myofibroblasts that promote profibrotic ECM synthesis; on the other hand, soft substrates prevent ECM synthesis. [
5,
15,
17,
41].
2. Hippo Signaling Pathway in Cardiac Fibrosis
The fibrotic response is a crucial contributor to heart failure (HF), which occurs in many types of cardiac diseases, including cardiac ischemia, myocardial infarction (MI), and cardiac hypertrophy. Understanding the mechanisms underlying fibrotic cardiac remodeling after injury remains a critical barrier to developing effective treatments for HF patients. An increasing number of studies have revealed important roles for Hippo signaling components in the development and progression of cardiac fibrosis [
18,
19,
20,
23,
27,
28,
29,
35,
37,
39,
42,
43,
44,
45,
46,
47,
48]. While YAP/TAZ activation is crucial for driving cardiac fibrosis, their suppression was beneficial for preventing angiotensin II (AngII) or MI-induced fibrosis [
18,
19,
29,
44].
Cardiac fibroblasts are believed to be the major source of pathological ECM synthesis during cardiac remodeling. Elevated YAP and downregulated upstream Hippo kinase LATS1 were determined in the left ventricular tissue of HF patients, which was associated with cardiac fibroblast proliferation [
42]. Recent studies also identified the activation of YAP and TAZ in resident cardiac fibroblasts from preclinical MI models (
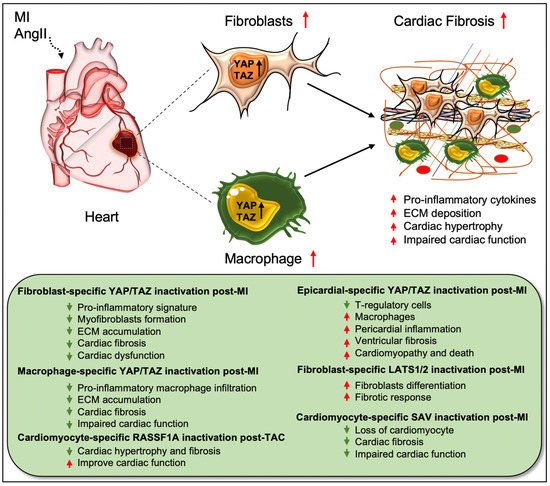
Figure 2) [
18,
19,
27]. YAP and TAZ can directly induce the differentiation of fibroblasts into pathologic myofibroblasts [
27,
28]. Fibroblast-specific deletion of
Yap/Taz using
Col1a2Cre(ER)T mice [
18] or deletion of only
Yap using
Tcf21MCM;
YapF/F mice [
19] showed reduced fibrotic and inflammatory responses in infarcted hearts. Similarly,
Yap/Taz deletion resulted in an impaired profibrotic response in the fibroblasts from the infarcted heart. Fibroblast-specific
Yap/Taz deficiency resulted in improved cardiac function in mice post-MI [
18]. Loss of YAP also reduced myocardial fibrosis and cardiac dysfunction in response to chronic neuroendocrine stimulation by AngII. In vitro studies revealed that blocking of YAP/TAZ with siRNA or verteporfin abrogated TGF-β1-induced transition of fibroblasts into myofibroblasts and ECM (such as collagen and fibronectin) production [
18]. In contrast, fibroblast-specific YAP overexpression using YAP
5SA mice promoted inflammatory response, fibrosis, and hypertrophy in the infarcted heart [
18]. Consistently, Francisco J et al. [
28] described that targeted overexpression of YAP in cardiac fibroblasts with adeno-associated virus construct AAV-hTCF21-FLAG-Yap(S127A)-augmented myocardial inflammation and fibrosis in mice, resulting in declined cardiac function. Mechanistically, ras homolog gene family member A (RhoA) regulates AngII-induced YAP activation, which mediates the transition of cardiac fibroblasts into fibrotic myofibroblasts [
19]. Moreover, YAP interacts with MRTF-A (myocardin-related transcription factor A) to facilitate the formation of α-smooth muscle actin (α-SMA)-positive myofibroblasts and profibrotic gene expression [
19]. YAP/TAZ also regulate interleukin-33 (IL-33) to promote cardiac myofibroblasts formation [
18]. These studies suggest that YAP and TAZ are crucially involved in regulating the formation of cardiac myofibroblasts and fibrosis.
Figure 2. Role of Hippo/YAP pathway in cardiac fibrosis and repair. Activation of YAP/TAZ in fibroblasts and macrophages plays a crucial role in driving cardiac fibrosis after MI or AngII-induced injury. Fibroblast-specific loss of Yap/Taz or loss of only Yap improves cardiac outcome and reduces fibrotic response in infarcted hearts post-injury. Genetic ablation of Yap/Taz in fibroblasts blocks the proliferation of fibroblasts and their differentiation into pathologic myofibroblasts and disrupts macrophage-mediated inflammatory signature. In contrast, fibroblast-specific inactivation of upstream Hippo kinases Lats1/2 aggravates fibrotic response post-MI. Like fibroblasts, conditional ablation of Yap/Taz in macrophages leads to impairment of macrophage-mediated inflammatory response and thereby reduces cardiac fibrosis, resulting in improved cardiac function and hypertrophy post-MI. Consistently, cardiomyocyte-specific inactivation of Hippo kinase Sav reduces cardiomyocyte apoptosis and cardiac fibrosis, which causes improved cardiac function post-MI. Likewise, cardiomyocyte-specific deletion of Rassf1A reduces TAC-induced cardiac hypertrophy and fibrosis with improved cardiac function. Epicardium-specific deficiency of Yap/Taz reduces the infiltration of T-regulatory cells (a subclass of adaptive immune cells) while increasing the recruitment of macrophages (a subclass of innate immune cells) in the myocardium, which promotes pericardial inflammation and ventricular fibrosis after MI-injury that subsequently leads to cardiomyopathy and death.
Xiao et al. [
35] elucidated that fibroblast-specific inactivation of upstream Hippo kinases LATS1/2 using
Tcf21iCre;
Lats1fl/fl;
Lats2fl/fl;
Rosa26mTmG mice initiated a spontaneous fibrotic response in the adult heart, which was aggravated after MI injury. The study also showed that LATS1/2 inhibits the differentiation of resting cardiac fibroblasts to myofibroblast-like cells. At the molecular level,
Lats1/2 mutant-cardiac fibroblasts showed reduced fibrotic phenotype due to reduced YAP/TAZ expression after infarction [
35]. Systemic loss of
Lats2 in mice alleviated myocardial fibrosis after transverse aortic constriction (TAC)-induced injury [
37]. Likewise, Lats1/2 inactivation in cardiomyocytes was also found to be protective against fibrosis after MI. Using
αMHCMerCreMer;
Savf/f mice, Leach et al. [
20] showed that
Sav deletion in cardiomyocytes improved cardiac function and reduced cardiac fibrosis with an increased presence of left ventricular cardiomyocytes at 9 weeks post-MI. Del Re et al. [
47] studied the crucial role of RASSF1A/MST1 pathway in TAC-induced cardiac fibrosis. RASSF1A is a physiological regulator of MST1 activation and the function of the RASSF1A/MST1 pathway is different between cardiomyocytes and fibroblasts. Overexpression of RASSF1A using
Rassf1A transgenic mice increased MST1 phosphorylation, cardiomyocyte apoptosis as well as cardiac fibrosis upon pressure overload. Alternately, mutations in
Rassf1A alleviated MST1 activation and improved cardiac function. Interestingly, systemic inactivation of
Rassf1A also aggravated the fibrotic response following TAC in the heart. The authors observed that cardiomyocyte-specific ablation of
Rassf1A, using
α-MHCCre/+ mice, attenuated TAC-induced cardiomyocyte apoptosis, cardiac hypertrophy, and fibrosis, and led to improvement in cardiac function. Interestingly, silencing of
Rassf1A in cardiac fibroblasts stimulated the TNFα-mediated proliferation of myofibroblasts and induced cardiac hypertrophy after pressure overload. These findings reveal cell-type-specific functions of RASSF1A after TAC-induced cardiac injury; inactivation of
Rassf1A in cardiomyocytes has protective effects while its inactivation in cardiac fibroblasts is detrimental to the heart.
Recent research has also described the role of Hippo mediators in cells other than fibroblasts and cardiomyocytes. For instance, Ramjee et al. [
46] demonstrated an epicardium-specific role of YAP/TAZ during myocardial recovery post-MI. Deletion of
Yap/Taz in the epicardium, using
Wt1CreERT2/+ mice, aggravated pericardial inflammation and ventricular fibrosis post-MI due to defective recruitment of regulatory T (Treg) cells (a subgroup of CD
4+ T cells) into the injured myocardium, resulting in cardiomyopathy and death. As a causal link, the study team found that YAP/TAZ regulates IFN-γ activity in the activated epicardium, which is important for myocardial recovery after MI. Epicardium-specific inactivation of
Yap/Taz caused reduced IFN-γ expression; the exogenous administration of IFN-γ post-MI by using a hydrogel system enhanced Treg cell recruitment into the injured myocardium and reduced fibrosis [
46]. Similarly, another recent study revealed a macrophage-specific role for YAP/TAZ in myocardial fibrosis, where the proteins were shown to function as essential regulators of macrophage-mediated proinflammatory or reparative responses post-MI in the heart [
29]. Conditional loss of
Yap/Taz, using
LysMCre mice, decreased pro-inflammatory and improved reparative macrophage polarization in the infarcted heart, leading to reduced cardiac fibrosis as well as improved cardiac outcomes. In contrast,
Yap overexpression, using a constitutively-active YAP mutant (YAP
5SA) and
LysMCre mice, caused an increase in inflammatory macrophage polarization and impairment in reparative macrophage polarization. As a consequence, YAP overexpression leads to exacerbated cardiac fibrosis and defective remodeling in the infarcted heart. These findings indicate that YAP/TAZ regulate post-MI macrophage polarization, which influences cardiac fibrosis and repair processes and, thereby, cardiac function (
Figure 2) [
29]. Since epicardium-specific ablation of
Yap/Taz-aggravated fibrosis and myeloid-specific inactivation reduced cardiac fibrosis and improved cardiac outcomes, targeted depletion of YAP/TAZ would be essential to prevent cardiac fibrosis post-MI.
Research over the years has revealed several components that have protective roles against cardiac fibrosis. For example, treatment of mice with MST1/2 blocker XMU-MP-1 prevented TAC-induced cardiomyocyte apoptosis, cardiac hypertrophy, and fibrosis [
39]. Similarly,
Qishen granules were shown to reduce the expression of fibrogenic proteins TGF-
β1, SMAD3, and CTGF in rats with HF [
43]. SKI, the cellular homolog of the avian Sloan-Kettering virus, negatively regulated the activation of primary rat cardiac fibroblasts by inducing proteasomal degradation of TAZ through interactions with LIM Domain-containing protein 1 (LIMD1) [
27]. Furthermore, blocking of RhoA attenuated AngII-induced YAP/TAZ transcriptional activity [
19] and suppression of YAP/TAZ signaling in mice by lovastatin alleviated AngII-induced fibrosis, both in vitro and in vivo (
Table 1) [
44]. The gaseous signaling molecule sulfur dioxide (SO2) efficiently prevented myocardial fibrosis by downregulating Hippo kinase MST1/2 in diabetic rats [
45].
This entry is adapted from the peer-reviewed paper 10.3390/cells11132065