Tissue hypoxia is one of the main pathophysiologic mechanisms in sepsis and particularly in COVID-19. Microvascular dysfunction, endothelialitis and alterations in red blood cell hemorheology are all implicated in severe COVID-19 hypoxia and multiorgan dysfunction. Tissue hypoxia results in tissue injury and remodeling with re-emergence of fetal programming via hypoxia-inducible factor-1α (HIF-1a)-dependent and -independent pathways. In this context, thyroid hormone (TH), a critical regulator of organ maturation, may be of relevance in preventing fetal-like hypoxia-induced remodeling in COVID-19 sepsis. Acute triiodothyronine (T3) treatment can prevent cardiac remodeling and improve recovery of function in clinical settings of hypoxic injury as acute myocardial infarction and by-pass cardiac surgery. Furthermore, T3 administration prevents tissue hypoxia in experimental sepsis. On the basis of this evidence, the use of T3 treatment was proposed for ICU (Intensive Care Unit) COVID-19 patients (Thy-Support, NCT04348513).
- thyroid hormone
- COVID-19
- erythrocyte
- sepsis
- right ventricle
- hypoxia
1. Introduction
2. Tissue Hypoxia: A Unifying Pathophysiologic Mechanism in Myocardial Infarction, Sepsis and Severe COVID-19
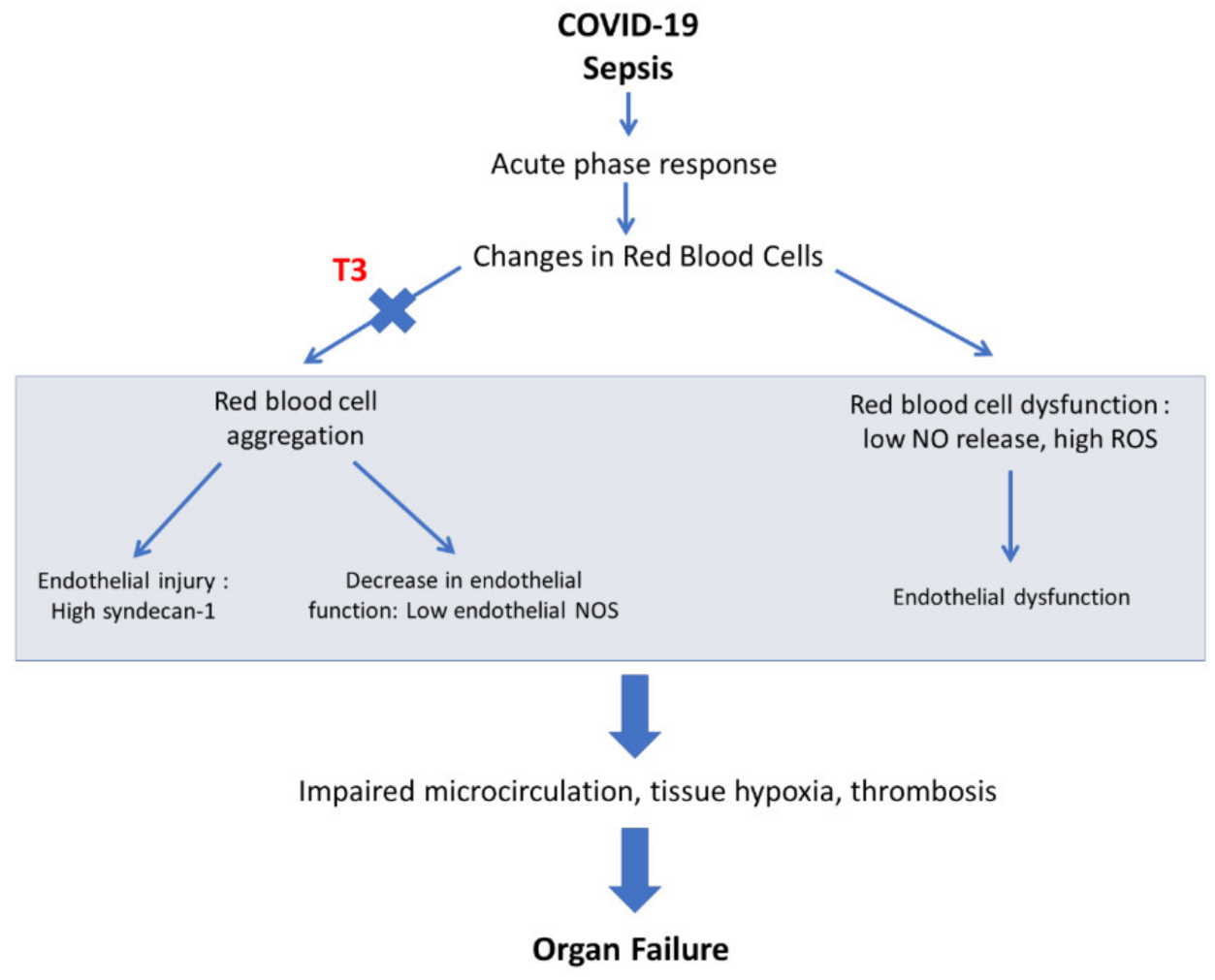
3. TH and Tissue Hypoxia
3.1. Differential Effects of TH on Healthy and Injured Tissue
3.2. Potential Underlying Mechanisms of TH Effects on Tissue Hypoxia
3.3. TH as Potential Therapy for Tissue Hypoxia
4. The Role of TH in COVID-19
This entry is adapted from the peer-reviewed paper 10.3390/ijerph19138063
References
- Padhi, R.; Kabi, S.; Panda, B.N.; Jagati, S. Prognostic significance of nonthyroidal illness syndrome in critically ill adult patients with sepsis. Int. J. Crit. Illn. Inj. Sci. 2018, 8, 165–172.
- Chen, W.; Tian, Y.; Li, Z.; Zhu, J.; Wei, T.; Lei, J. Potential Interaction Between SARS-CoV-2 and Thyroid: A Review. Endocrinology 2021, 162, bqab004.
- Johansson, P.I.; Stensballe, J.; Ostrowski, S.R. Shock induced endotheliopathy (SHINE) in acute critical illness—A unifying pathophysiologic mechanism. Crit. Care 2017, 21, 25.
- Pantos, C.; Mourouzis, I.; Cokkinos, D.V. Rebuilding the post-infarcted myocardium by activating ‘physiologic’ hypertrophic signaling pathways: The thyroid hormone paradigm. Heart Fail. Rev. 2010, 15, 143–154.
- Pantos, C.; Mourouzis, I. Thyroid hormone receptor α1 as a novel therapeutic target for tissue repair. Ann. Transl. Med. 2018, 6, 254.
- Pantos, C.I.; Trikas, A.G.; Pisimisis, E.G.; Grigoriou, K.P.; Stougiannos, P.N.; Dimopoulos, A.K.; Linardakis, S.I.; Alexopoulos, N.A.; Evdoridis, C.G.; Gavrielatos, G.D.; et al. Effects of acute triiodothyronine treatment in patients with anterior myocardial infarction undergoing primary angioplasty: Evidence from a pilot randomized clinical trial (ThyRepair study). Thyroid 2022, 32, 714–724.
- Mourouzis, I.S.; Lourbopoulos, A.I.; Trikas, A.G.; Tseti, I.K.; Pantos, C.I. Triiodothyronine prevents tissue hypoxia in experimental sepsis: Potential therapeutic implications. Intensive Care Med. Exp. 2021, 9, 17.
- Dekker, N.A.M.; Veerhoek, D.; Koning, N.J.; van Leeuwen, A.L.I.; Elbers, P.W.G.; van den Brom, C.E.; Vonk, A.B.A.; Boer, C. Postoperative microcirculatory perfusion and endothelial glycocalyx shedding following cardiac surgery with cardiopulmonary bypass. Anaesthesia 2019, 74, 609–618.
- Vincent, J.L.; De Backer, D. Microvascular dysfunction as a cause of organdysfunction in severe sepsis. Crit. Care 2005, 9 (Suppl. S4), S9–S12.
- Sakr, Y.; Dubois, M.J.; De Backer, D.; Creteur, J.; Vincent, J.L. Persistent microcirculatory alterations are associated with organ failure and death in patients with septic shock. Crit. Care Med. 2004, 32, 1825–1831.
- Konijnenberg, L.S.F.; Damman, P.; Duncker, D.J.; Kloner, R.A.; Nijveldt, R.; van Geuns, R.M.; Berry, C.; Riksen, N.P.; Escaned, J.; van Royen, N. Pathophysiology and diagnosis of coronary microvascular dysfunction in ST-elevation myocardial infarction. Cardiovasc. Res. 2020, 116, 787–805.
- van Kranenburg, M.; Magro, M.; Thiele, H.; de Waha, S.; Eitel, I.; Cochet, A.; Cottin, Y.; Atar, D.; Buser, P.; Wu, E.; et al. Prognostic value of microvascular obstruction and infarct size, as measured by CMR in STEMI patients. JACC Cardiovasc. Imaging 2014, 7, 930–939.
- Kanoore Edul, V.S.; Caminos Eguillor, J.F.; Ferrara, G.; Estenssoro, E.; Siles, D.S.P.; Cesio, C.E.; Dubin, A. Microcirculation alterations in severe COVID-19 pneumonia. J. Crit. Care 2021, 61, 73–75.
- Sloop, G.D.; Pop, G.A.; Weidman, J.J.; Cyr, J.A. The Detrimental Role of Elevated Blood Viscosity in Patients With COVID-19. J. Cardiol. Ther. 2021, 8, 976–980.
- Johansson, P.I.; Stensballe, J.; Rasmussen, L.S.; Ostrowski, S.R. A high admission syndecan-1 level, a marker of endothelial glycocalyx degradation, is associated with inflammation, protein C depletion, fibrinolysis, and increased mortality in trauma patients. Ann. Surg. 2011, 254, 194–200.
- Johansen, M.E.; Johansson, P.I.; Ostrowski, S.R.; Bestle, M.H.; Hein, L.; Jensen, A.L.; Soe-Jensen, P.; Andersen, M.H.; Steensen, M.; Mohr, T.; et al. Profound endothelial damage predicts impending organ failure and death in sepsis. Semin. Thromb. Hemost. 2015, 41, 16–25.
- Zhang, D.; Li, L.; Chen, Y.; Ma, J.; Yang, Y.; Aodeng, S.; Cui, Q.; Wen, K.; Xiao, M.; Xie, J.; et al. Syndecan-1, an indicator of endothelial glycocalyx degradation, predicts outcome of patients admitted to an ICU with COVID-19. Mol. Med. 2021, 27, 151.
- Ackermann, M.; Verleden, S.E.; Kuehnel, M.; Haverich, A.; Welte, T.; Laenger, F.; Vanstapel, A.; Werlein, C.; Stark, H.; Tzankov, A.; et al. Pulmonary Vascular Endothelialitis, Thrombosis, and Angiogenesis in COVID-19. N. Engl. J. Med. 2020, 383, 120–128.
- Baskurt, O.K.; Yalcin, O.; Ozdem, S.; Armstrong, J.K.; Meiselman, H.J. Modulation of endothelial nitric oxide synthase expression by red blood cell aggregation. Am. J. Physiol. Heart Circ. Physiol. 2004, 286, H222–H229.
- Iffrig, E.; Oakley, J.; Fibben, K.; Sakurai, Y.; Esper, A.M.; Lam, W. Severe COVID-19 Is an Acquired Red Cell Disease- Potential Mechanisms Underlying Endothelialitis. Am. J. Respir. Crit. Care Med. 2021, 203, A4470.
- Baskurt, O.K.; Temiz, A.; Meiselman, H.J. Red blood cell aggregation in experimental sepsis. J. Lab. Clin. Med. 1997, 130, 183–190.
- Li, C.; Wang, Y.; Zhang, Q.; Wang, L.; Li, K.; Yang, X. Incorporating the erythrocyte sedimentation rate for enhanced accuracy of the global registry of acute coronary event score in patients with ST-segment elevated myocardial infarction: A retrospective cohort study. Medicine 2020, 99, e22523.
- Nader, E.; Nougier, C.; Boisson, C.; Poutrel, S.; Catella, J.; Martin, F.; Charvet, J.; Girard, S.; Havard-Guibert, S.; Martin, M.; et al. Increased blood viscosity and red blood cell aggregation in patients with COVID-19. Am. J. Hematol. 2022, 97, 283–292.
- Brugnara, C. The Erythrocyte, a Novel Disease-Mediator for COVID-19 Vasculopathy? JACC Basic Transl. Sci. 2022, 7 Pt 1, 205–206.
- Mahdi, A.; Collado, A.; Tengbom, J.; Jiao, T.; Wodaje, T.; Johansson, N.; Farnebo, F.; Färnert, A.; Yang, J.; Lundberg, J.O.; et al. Erythrocytes Induce Vascular Dysfunction in COVID-19. JACC Basic Transl. Sci. 2022, 7 Pt 1, 193–204.
- Lourbopoulos, A.I.; Mourouzis, I.S.; Trikas, A.G.; Tseti, I.K.; Pantos, C.I. Effects of Thyroid Hormone on Tissue Hypoxia: Relevance to Sepsis Therapy. J. Clin. Med. 2021, 10, 5855.
- Ferreyra, C.; O’Valle, F.; Osorio, J.M.; Moreno, J.M.; Rodriguez, I.; Vargas, F.; Osuna, A. Effect of preconditioning with triiodothyronine on renal ischemia/reperfusion injury and poly(ADP-ribose) polymerase expression in rats. Transpl. Proc. 2009, 41, 2073–2075.
- Sutter, P.M.; Thulin, G.; Stromski, M.; Ardito, T.; Gaudio, K.M.; Kashgarian, M.; Siegel, N.J. Beneficial effect of thyroxin in the treatment of ischemic acute renal failure. Pediatr. Nephrol. 1988, 2, 1–7.
- Ferreyra, C.; Vargas, F.; Rodriguez-Gomez, I.; Perez-Abud, R.; O’Valle, F.; Osuna, A. Preconditioning with triiodothyronine improves the clinical signs and acute tubular necrosis induced by ischemia/reperfusion in rats. PLoS ONE 2013, 8, e74960.
- Friederich-Persson, M.; Persson, P.; Fasching, A.; Hansell, P.; Nordquist, L.; Palm, F. Increased kidney metabolism as a pathway to kidney tissue hypoxia and damage: Effects of triiodothyronine and dinitrophenol in normoglycemic rats. Adv. Exp. Med. Biol. 2013, 789, 9–14.
- Sivertsson, E.; Friederich-Persson, M.; Persson, P.; Nangaku, M.; Hansell, P.; Palm, F. Thyroid hormone increases oxygen metabolism causing intrarenal tissue hypoxia; a pathway to kidney disease. PLoS ONE 2022, 17, e0264524.
- Geist, D.; Hones, G.S.; Gassen, J.; Kerp, H.; Kleinbongard, P.; Heusch, G.; Fuhrer, D.; Moeller, L.C. Noncanonical Thyroid Hormone Receptor alpha Action Mediates Arterial Vasodilation. Endocrinology 2021, 162, bqab099.
- Pantos, C.; Mourouzis, I.; Cokkinos, D.V. Thyroid hormone and cardiac repair/regeneration: From Prometheus myth to reality? Can. J. Physiol. Pharmacol. 2012, 90, 977–987.
- Forini, F.; Nicolini, G.; Iervasi, G. Mitochondria as key targets of cardioprotection in cardiac ischemic disease: Role of thyroid hormone triiodothyronine. Int. J. Mol. Sci. 2015, 16, 6312–6336.
- Pantos, C.I.; Malliopoulou, V.A.; Mourouzis, I.S.; Karamanoli, E.P.; Paizis, I.A.; Steimberg, N.; Varonos, D.D.; Cokkinos, D.V. Long-term thyroxine administration protects the heart in a pattern similar to ischemic preconditioning. Thyroid 2002, 12, 325–329.
- Pantos, C.; Mourouzis, I.; Saranteas, T.; Clave, G.; Ligeret, H.; Noack-Fraissignes, P.; Renard, P.Y.; Massonneau, M.; Perimenis, P.; Spanou, D.; et al. Thyroid hormone improves postischaemic recovery of function while limiting apoptosis: A new therapeutic approach to support hemodynamics in the setting of ischaemia-reperfusion? Basic Res. Cardiol. 2009, 104, 69–77.
- Pantos, C.; Mourouzis, I.; Saranteas, T.; Brozou, V.; Galanopoulos, G.; Kostopanagiotou, G.; Cokkinos, D.V. Acute T3 treatment protects the heart against ischemia-reperfusion injury via TRalpha1 receptor. Mol. Cell. Biochem. 2011, 353, 235–241.
- Pantos, C.; Mourouzis, I.; Galanopoulos, G.; Gavra, M.; Perimenis, P.; Spanou, D.; Cokkinos, D.V. Thyroid hormone receptor alpha1 downregulation in postischemic heart failure progression: The potential role of tissue hypothyroidism. Horm. Metab. Res. 2010, 42, 718–724.
- Mourouzis, I.; Mantzouratou, P.; Galanopoulos, G.; Kostakou, E.; Roukounakis, N.; Kokkinos, A.D.; Cokkinos, D.V.; Pantos, C. Dose-dependent effects of thyroid hormone on post-ischemic cardiac performance: Potential involvement of Akt and ERK signalings. Mol. Cell. Biochem. 2012, 363, 235–243.
- Ranasinghe, A.M.; Quinn, D.W.; Pagano, D.; Edwards, N.; Faroqui, M.; Graham, T.R.; Keogh, B.E.; Mascaro, J.; Riddington, D.W.; Rooney, S.J.; et al. Glucose-insulin-potassium and tri-iodothyronine individually improve hemodynamic performance and are associated with reduced troponin I release after on-pump coronary artery bypass grafting. Circulation 2006, 114 (Suppl. S1), I245–I250.
- Ntari, L.; Mantzouratou, P.; Katsaouni, A.; Pantos, C.; Kollias, G.; Mourouzis, I. Changes in Thyroid Hormone Signaling Mediate Cardiac Dysfunction in the Tg197 Mouse Model of Arthritis: Potential Therapeutic Implications. J. Clin. Med. 2021, 10, 5512.
- Pantos, C.; Xinaris, C.; Mourouzis, I.; Kokkinos, A.D.; Cokkinos, D.V. TNF-alpha administration in neonatal cardiomyocytes is associated with differential expression of thyroid hormone receptors: A response prevented by T3. Horm. Metab. Res. 2008, 40, 731–734.
- Baldelli, R.; Nicastri, E.; Petrosillo, N.; Marchioni, L.; Gubbiotti, A.; Sperduti, I.; Di Giacinto, P.; Rizza, L.; Rota, F.; Franco, M.; et al. Thyroid dysfunction in COVID-19 patients. J. Endocrinol. Investig. 2021, 44, 2735–2739.
- Lui, D.T.W.; Lee, C.H.; Chow, W.S.; Lee, A.C.H.; Tam, A.R.; Fong, C.H.Y.; Law, C.Y.; Leung, E.K.H.; To, K.K.W.; Tan, K.C.B.; et al. Thyroid Dysfunction in Relation to Immune Profile, Disease Status, and Outcome in 191 Patients with COVID-19. J. Clin. Endocrinol. Metab. 2021, 106, e926–e935.
- Lang, S.; Liu, Y.; Qu, X.; Lu, R.; Fu, W.; Zhang, W.; Wang, H.; Hong, T. Association between Thyroid Function and Prognosis of COVID-19: A Retrospective Observational Study. Endocr. Res. 2021, 46, 170–177.
- Ballesteros Vizoso, M.A.; Castilla, A.F.; Barceló, A.; Raurich, J.M.; Argente del Castillo, P.; Morell-García, D.; Velasco, J.; Pérez-Bárcena, J.; Llompart-Pou, J.A. Thyroid Disfunction in Critically Ill COVID-19 Patients. Relationship with In-Hospital Mortality. J. Clin. Med. 2021, 10, 5057.
- Chen, Y.; Li, X.; Dai, Y.; Zhang, J. The Association Between COVID-19 and Thyroxine Levels: A Meta-Analysis. Front. Endocrinol. 2021, 12, 779692.
- Muller, I.; Cannavaro, D.; Dazzi, D.; Covelli, D.; Mantovani, G.; Muscatello, A.; Ferrante, E.; Orsi, E.; Resi, V.; Longari, V.; et al. SARS-CoV-2-related atypical thyroiditis. Lancet Diabetes Endocrinol. 2020, 8, 739–741.
- Rotondi, M.; Coperchini, F.; Ricci, G.; Denegri, M.; Croce, L.; Ngnitejeu, S.T.; Villani, L.; Magri, F.; Latrofa, F.; Chiovato, L. Detection of SARS-CoV-2 receptor ACE-2 mRNA in thyroid cells: A clue for COVID-19-related subacute thyroiditis. J. Endocrinol. Investig. 2021, 44, 1085–1090.
- Samuels, M.H.; McDaniel, P.A. Thyrotropin levels during hydrocortisone infusions that mimic fasting-induced cortisol elevations: A clinical research center study. J. Clin. Endocrinol. Metab. 1997, 82, 3700–3704.
- Nicoloff, J.T.; Fisher, D.A.; Appleman, M.D., Jr. The role of glucocorticoids in the regulation of thyroid function in man. J. Clin. Investig. 1970, 49, 1922–1929.