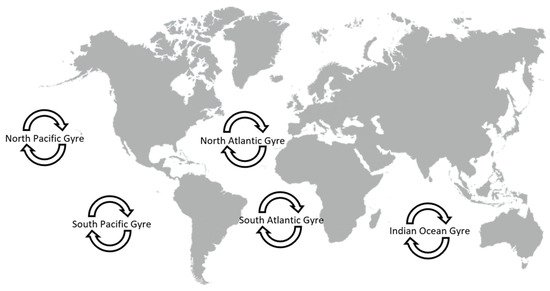
4. Engineered Nanoparticles
Particles of up to 100 nm in diameter are known as nanoparticles. The European Union defines a suspension of nanoparticles as that in which 50% or more particles have one or more external dimensions of 1–100 nm. These suspensions exist in nature (e.g., nanoclays), but in the recent past, they have been manufactured for specific human purposes. Such engineered nanoparticles (ENPs) may be used, for example, in coatings, insulating and magnetic materials, and as antimicrobial additives, principally nanometal oxides, such as TiO
2, which become increasingly antimicrobial under the action of UV In fact, metal and metal oxide nanoparticles are those most produced worldwide, with TiO
2 having the highest produced mass [
171], and many, along with silica nanoparticles, are used as antifouling materials for protection of marine and seawater-associated structures [
172,
173,
174]. The antifouling materials can, of course, also affect the non-target biota in the sea, especially when liberated from the protected structures by sloughing or friction. Toxic effects of the principal metal oxide ENPs in the marine environment decrease in the order Au > Zn > Ag > Cu > Ti > Carbon60 [
175].
At some stage in their lifetime, even if not utilized directly in the marine environment, ENPs will be released into their surroundings and end up entering the oceans, where they can exert negative effects on the biota [
176,
177], including inhibition of movement and metabolic processes, oxidative stress and dysfunctional DNA replication [
178]. NanoZnO particles ranging from 30 nm to 2 μm have been shown to be highly toxic to flounder cells in culture and zebrafish embryos [
179]. ENPs can readily pass from terrestrial waters into the ocean and thence into the marine food web [
180]. It has been suggested that the main effects of ENPs on coastal marine life forms will be in sediments [
181], where they have been shown to have toxic effects on foraminifera [
182].
There has been little empirical study of the fate of ENPs in the aquatic environment [
183], although it has been suggested that they can cause significant harm to the marine ecosystem [
184] and references therein and that they have significant toxic effects on marine phytoplankton [
185]. The release of silver NPs has been shown to alter the functioning of the marine food web by hampering important viral and bacterial processes [
186]. In a mesocosm experiment, the addition of silver NPs, even at a low dose, affected planktonic communities, especially reducing the growth of the cyanobacterium
Synechococcus. Viral auxiliary metabolic genes involved in cyanobacterial photosynthesis were also decreased.
It has, however, been suggested recently that ENPs may not be found at sufficiently high concentrations in the natural environment to pose a current problem [
187]. More data on the effects and fate of nanoparticles released into the environment are necessary. Life cycle and ecological risk assessments of ENPs in our oceans are essential to stimulate remediation processes and protect the marine environment.
5. Metallic Particles
Mining of polymetallic nodules, found on the surface of abyssal plains at around 4000 m depth, results in the release into the benthos of sediment plumes and nodule debris. These can be rich in Mn, Ni, Cu, and Co [
188]. Fazey and Ryan [
164] examined the aerobically grown bacteria present on the surface of nodules and in the overlying sediment, identifying
Halomonas aquamarina,
H. meridiana, and
Erythrobacter citreus in both, but the genera
Arthrobacter,
Kocuria,
Loktanella,
Marinobacter and
Pseudoalteromonas only in sediment within 4 cm of the nodule surface. Cho et al. [
189] confirmed that the microbiome of nodules differs from the microbial population in the surrounding sediment, but their use of NGS technology led to the detection of a different set of bacteria and Archaea. Thaumarchaeota were found in both sediment and nodule, Mn-oxidizing bacteria (
Hyphomycrobium, Aurantimonas, and
Marinobacter) were predominant in nodules, and
Idiomarina,
Erythrobacter, and
Sulfitobacter in sediments. Gillard et al. [
188], based on their analyses, suggest the use of standard cultivation techniques for monitoring plume propagation; indicator organisms for sediment would be
Diezia maris and
Pseudoalteromonas shioyasakiensis, for nodules
Rhodococcus erythropolis and water
Marinobacter flavimaris.
Much of the iron found in aerosols over the oceans is anthropogenic in origin, resulting from the burning of fossil and biofuels and fires on land (biomass burning), a situation that is probably mirrored by zinc [
190]. Conway et al. [
191], using iron-isotope ratios, showed that deposition of anthropogenic Fe could reach almost 100% of the total Fe near highly populated areas. Their model suggested that this effect would be greatest in the Southern and Pacific Oceans, and this was echoed by Hamilton et al. in 2020 [
192]. Much of the iron, and, indeed, many metals found in marine particles may be linked to the presence of microorganisms that produce metal-chelating siderophores. Chuang et al. [
193] found that hydroxamate siderophores comprised a large part of the sinking particles (“marine snow” q.v.) collected in the Sargasso Sea. One of the important ecological functions of the siderophores produced by microorganisms is the release of iron from sinking particles to supply dissolved iron to the water column [
194]. The export of iron from hydrothermal vents in the Southern East Pacific has likewise been linked to particles containing microorganisms [
195].
Pollution by mercuric ions is a potential risk to human health, principally through the consumption of fish [
196]. The main anthropogenic source of this metal is artisanal and small-scale gold mining, followed by the burning of fossil fuels [
197]. The metal is converted to toxic methylmercury and dimethylmercury by microbial activity in the seas [
198] and is largely associated with marine particulate matter [
199]. The latter authors identified the sulfate-reducing bacterium,
Desulfovibrio desulfuricans, as important for the uptake or exchange of Hg
2+ in anaerobic environments. Marine Group II (MGII) archaeal genes associated with assimilatory sulfate reduction have been detected, along with MGII genes involved in surface adhesion, in samples collected from around the world during the Tara Oceans’ circumnavigation trip [
156]. The authors suggested that archaea MGII could be implicated in the degradation of marine particles, a more positive role for microbial biofilms in our oceans.
Marine microorganisms are also important in the production of metallic compounds. The mineral barite (or baryte), used principally in drilling muds, is produced in the oceans by barium binding initially to phosphate groups in bacterial cells or EPS; the thus concentrated barium is then converted in the marine environment to barite [
200].
6. Sinking Particles (“Marine Snow”) and Pollution
Marine snow is considered to be composed of heterogeneous agglomerates of living and dead organic matter of >500 um in size, formed by the attachment of organisms to the so-called “transparent exopolymer particles” (TEPs) that consist mainly of acidic polysaccharides previously produced by phytoplankton and heterotrophic prokaryotes [
201] and references therein. The particles contain diverse groups of eukaryotes, which may somewhat resemble, but certainly do not equal, the plankton in the local environment [
202]. They have a highly variable composition and there are fundamental differences in particle composition between oligotrophic and eutrophic environments [
202]. The microbial taxa associated with the particles are very different from those in the surrounding seawater and may contain increased oil degraders in oil-polluted environments [
203,
204] or methylmercury genes in saline waters in the North Sea [
205]. Hence the particles may be “hot spots” for the degradative activity of surrounding pollutants [
206].
Such sinking particles differ from floating particles in the oceans in carrying a changing population of prokaryotic species. Duret et al. [
207] identified the prokaryotic populations on both types of particles in the Scotia Sea (Southern Ocean) and suggested that
r-strategists, with generalized metabolic activities and rapid substrate consumption, were better adapted to sinking particles, with their changing environment, while
K-strategists, specialized for complex organic material degradation, were better adapted to the more stable environment of semi-labile floating particles. So, for instance, pseudomonads and Rhodobacteriales were enriched on sinking particles, Flavobacteriales on floating. Datta et al. [
208] had previously shown, using model polysaccharide particles, that the attached bacterial communities underwent rapid metabolic successions, driven by the environment. They suggested that there are 3 phases of colonization: attachment of a highly diverse community, selection of specific metabolic activities by the environment (reducing diversity) and replacement by secondary consumers, metabolizing the products of the second phase cells, and increasing diversity somewhat once more. Liu et al. [
209], investigating differences between the two types of particles at low and high pressures in the New Britain Trench, Solomon Sea (Pacific), found that, although there were differences in prokaryotic populations on floating and sinking particles, similar groups participated in the degradation of diatom debris.
Even if similar organisms are involved in degradation, the physical act of sinking, whereby water flows past the particle surface, increases the rate of biodegradation simply by aiding the removal of the degradation products, driving the reaction to the right. This increase in microbiodegradation in sinking, as opposed to static, particles was elegantly demonstrated in a mathematical model developed by Alcolombri et al. [
210].
The presence of eukaryotes in marine snow has been less frequently investigated. Bochdansky et al. [
211] showed the presence of a fungal biomass equal to that of prokaryotes in bathypelagic particles from the North Atlantic and Arctic seas. Fungi and labyrinthulomycetes (the latter mainly labyrinthulids and thraustochytrids) dominated the biomass. These eukaryotes are tolerant of low temperatures and high pressures and were considered to be potentially important biodegraders in the particles. Schultz et al. [
212], using metaproteomics and functional analyses of marine particles, showed that eukaryotes were more abundant in the particles than in the surrounding seawater. Those detected in particles were phytoplankton, Oomycetes, and Fungi. Greater amounts of viral proteins were also found in the particles. They reported rather small differences between bacterial proteins on particles and in the planktonic phase. The relative abundance of eukaryotes and viruses confirmed the results of López-Pérez et al. [
213], who investigated the coastal waters off Alicante, Spain. They also found that there was an overrepresentation in the particle-associated microbiome of alpha, delta, and gamma proteobacteria, bacteroidetes (Flavobacteria), Planktomycetes, and Actinobacteria.