The human brain is one of the most dynamically intricate networks molded by nature capable of performing a wide array of activities effectively and efficiently [
1,
2,
3,
4]. Operating on a high degree of complexity, brain dynamics consist of rapid reconfiguration of network states driven by interactions between network constituents to optimize temporal global evolution [
5,
6]. Constituents from the micro to the macro scale, such as neural cells, cluster to brain nuclei, and regions interplay with one another to compose an instantaneous, dynamical form of the brain, which serves to interact with the environment [
7,
8]. Brain dynamics are unified across its spatiotemporal scales to work in concert to coordinate an instantaneous current representation while simultaneously maintaining active recollections and processing of prior experiences, along with evolutionary developed, primal, raw, emotional contexts, which can influence future trajectories and goals for the brain [
9,
10]. Constituent parts or subsystems of a network have unique responsibilities in contributing towards the overall time evolution of a network [
11,
12]. Thus, components of the brain cooperate and, in some cases, compete with one another from the micro to macro scales to direct and determine temporal evolution of the network’s global behaviors [
13]. Examples of these include neocortical modulation of amygdala activity to initiate higher-order cognitive regulation upon potentially fearful stimuli [
14]. This interaction illustrates how activity produced by limbic regions (amygdala and associated areas), which provide primal emotional motivations such as fear, is regulated by contributions from the neocortex, which provides more complex forms of information manipulation, rendering higher cognitive thought to assess the initial appraisals of emotional response (such as fear) with more logic [
15]. Furthermore, local activity from these regions are routed to one another via the thalamus, a relay center in the brain capable of coupling neocortical activity with a variety of localized subcortical structures. The resulting collaboration (or competition), sways global network trajectory towards a particular path [
16]. The brain must simultaneously organize and process these various modes of information to construct an instinctual network system reaction, ensuring coherent brain behavior. Information is physically transmitted via configured patterns of electrophysiological neural activity. Upon accomplishing this, the brain can contextualize its network state within the time-varying environment. Learning from previous experiences, executing current actions, and preparing future expectations consists of these dynamical capabilities, enabling the brain to optimize the variety of possible opportunities posed by the the time-varying environment, ranging from scavenging food to maneuvering social situations and assessing potential sexual partners.
Brain dynamics are defined as the global neural processes that direct the network’s evolution in time, commonly seen and experienced by the processing of sensory input and formulating a corresponding output [
38]. These are typically observed in the change of the characteristics of the brain seen in the time-varying properties of the cumulative neuronal assemblies [
39]. Experimental approaches observe this in the electrical activity of groups of neurons through electroencephalography (EEG) measurements or blood flow across brain regions through blood oxygen level dependency (BOLD) analysis via functional magnetic resonance imaging (fMRI) and how these properties change with exposure to new input [
40,
41]. It must be noted that these methods do not explicitly isolate component neuronal activity. For example, fMRI detects changes in blood flow related to brain activity (formally described as BOLD analysis). Naturally, as the brain evolves over time, resources are redistributed by altering blood flow, which is detectable through fMRI; however, the resolution of this observable change is not sufficient to delineate the firing properties and patterns down to the scale of individual neurons. In addition to limitations of spatial resolution, fMRI-centered BOLD analysis lacks the temporal resolution to identify the time evolution of a neural component’s firing patterns at the millisecond scale [
42]. On the other hand, it is also difficult to isolate component neural activity at sufficient resolutions using EEG, as the detected EEG waveform is a superposition of dynamic electromagnetic activity, including local field potentials generated through the cumulative ionic flux in and out of the cellular space [
43]. Additional techniques using magnetoencephalography (MEG) detect changes in magnetic fields resulting from dynamic electrical currents produced in the brain from neuronal activities. These represent examples of observed changes in brain structure and function [
44]. The interpretations of these methods have been refined over the years with the addition of advanced techniques [
45,
46]. Whereas concrete claims remain elusive due to a lack of temporal or spatial resolution, a commonly observed theme is that there is no stationary state of the brain [
47]. For example, classical EEG experiments have framed brains as nonequilibrium systems along with the observation that unique patterns of EEG waveforms acquired from the olfactory bulb correspond towards information processing of specific odors [
48]. These established studies make it apparent that the brain does not remain in a static configuration; its form changes to varying degrees over time. Therefore, the brain is fundamentally a nonstationary system without an equilibrium point that utilizes its biological capabilities to detect, interpret, and respond to the dynamical environment. Portions of this complexity are apparent through observable neurodynamic rhythms seen in EEG or fMRI recordings. Despite this recognition, the exact underpinnings of this substantial degree of complexity are among the core questions, ambiguities, and mysteries of modern neuroscience.
It must be recognized that significant understanding has been achieved through the earliest developments in neuroscience accomplished by Cajal and Broca, along with more recent undertakings utilizing the tools developed in network sciences, which have contributed to the development of a transdisciplinary perspective. Neuroscience research has been traditionally led by animal models, advanced neuroimaging techniques, brain tissue sampling, and separation methods [
49,
50,
51]. These procedures have generated notable accomplishments, such as having a fundamental knowledge in identifying neuronal cell-mechanisms, structures, and functions, including dendritic and synaptic regulation, to identify and classify individuals, connections, and populations of neurons. Conventional approaches in neuroscience have led this progress; however, a comprehensive understanding of brain dynamical phenomena is still lacking in terms of how local and global cognitive mechanisms interplay simultaneously across multivariate scales. A transdisciplinary field of network sciences has emerged over the past 20 years in attempts to address complexity in the brain and other complex networks and has met with limited success, particularly in helping to realize that a transdisciplinary perspective is necessary to guide the next level of progress in neuroscience [
52]. A brief review of the merits and limits of network sciences follows. Traditional network science has been spearheaded by graph theory, defining individuals in a network as nodes and their interactions as edgewise connections between nodes [
53]. It is important to note that this is purely a mathematically driven formalism that is not necessarily driven by fundamental physical law. Small-world and scale-free network models have influenced the development of established network theories over the past 20 years [
54,
55]. For example, graph theory developments have been used to topologically describe networks and have been translated into anatomical and functional brain networks [
56]. These are suited to capture small-world topology, such as highly interconnected hubs and modularity prevalent in the brain [
57]. Additional topological properties of complex networks, such as hierarchies, centrality, and network hub distribution, have also been realized in this process [
58]. Using serial reconstructions of electron microscopy, a complete connection matrix of the nematode C. elegans has been accomplished and described as a small-world network [
59]. Furthermore, using combinations of physiological and anatomical techniques, multielectrode activity recordings have generated reconstructions of cellular networks in the neocortex, and diffusion tensor imaging has developed a map for cortical and basal brain gray matter areas [
60]. The interplay of these methods has inspired a plethora of studies, models, and reviews [
61,
62,
63]. These archetypes represent characteristics observed in networks under limitations. The assumptions underlying these limitations for small-world and scale-free networks must be considered when determining real-world applicability. For example, the network description is time-invariant, which neglects the dynamical elements inherent in all complex networks. Misrepresenting the dynamics can lower the accuracy of analysis at best or lead to catastrophic failure at worst. If the local interactions in a network are static, the global dynamics are adulterated and insufficient. Temporal networks are developed in attempts to compensate for this [
64]. These models help represent the time-varying qualities of network structures, such as multilayer dynamics [
65,
66]. Whereas these help in developing tools better geared towards the dynamical aspects of complex networks, many of these methods still are plagued with the limited applicability of graph theory. For example, interactions represented by stationary edgewise connections between individuals lack the highly nonlinear features present in networks with diverse connections between individuals, groups, and large populations (composed of smaller groups and individuals) [
67]. Misrepresentation of this fundamental nonlinearity and dynamics renders traditional methods inept for comprehensive analysis and control. Additionally, a pure mathematical representation of a network ensures quantitative precision; however, the current state of network sciences does not necessarily intertwine this foundation with fundamental physical laws, compromising its comprehensive accuracy.
2. Nonlinear Biological Interactions
This section will express the nonlinear nature of local interactions and how these contribute towards global network properties. After this section, details on global network properties (including the form and structures of higher order neurodynamic complexity) will be reviewed in detail. For now, the global state of brain phenomena is a time-varying ensemble, consistently changing to different degrees in accordance with factors within and without. Thus, brain phenomena are consistently nonstationary to different degrees in accordance with different environmental perturbations navigated through nonlinear interactions, propelling a wide repertoire of dynamics [
68]. The properties of these local interactions determine global form and function. Therefore, to better understand the macroscopic brain, one must begin first with the brain’s auxiliary local interactions. As they cumulatively dictate global function, local interactions represent physical connections (or interactions) that deem the magnitude and direction of influence one agent has on another in a network and can be viewed as degrees of coupling [
69]. These local interactions between connecting agents, regions, and subnetworks in the brain allow smaller-scale subsystems to coordinate with one another, composing coherent global forms by promoting coordinated local interactions, which engender stable global brain dynamics [
70,
71]. Thus, dynamical overall brain activity is nurtured through flexible configuration of local connectivity capable of generating a diverse variety of brain behaviors [
72]. These include axonal architectures [
73] with adaptive myelination [
74], complex configurations of dendritic branching [
75] and dendritic spine morphology [
76], as well as the dynamic synapse [
77], housing a multitude of pre- and postsynaptic mechanisms [
78]. Importantly, each of these mechanisms is nonstationary and capable of dynamically influencing neural interactions along a wide range of spatiotemporal scales. Thus, local interactions range from (1) microscopic interactions between individual neurons and glial cells to (2) interplay between clusters of nuclei in the brain to (3) mesoscopic relations between different regions of the brain, to highlight a select few (out of the many scales in the brain). The cumulation of these interactions, along with others not mentioned or yet to be discovered, is built to fine-tune connections between local brain regions operating on a variety of temporal and spatial scales. Due to these diverse factors of coupling, which can change on a variety of time scales, interactions are fundamentally nonlinear in the time-domain. Furthermore, nonlinearity, observed in the dynamical interactions amongst a wide distribution of neural frequencies, engenders highly nonlinear characteristics simultaneously in the frequency domain. Moving forward, these produce highly nonlinear characteristics in overall spatiotemporal brain dynamics, enabling the unprecedented level of network reconfiguration observed and experienced in the human brain. Thus, the simultaneous nonlinearity in the time and frequency domains elicits signature characteristics of chaos, which are essential for rapid reconfiguration of brain network states [
79]. This topic is worthy of a detailed discussion for another review; however, for the context of this paper, it must be borne in mind that the level of global complexity in the brain is a product of its local nonlinearities at the fundamental level. In other words, the flexible nature of the connections (interactions) between individual parts of a brain network across its many scales and modes of operation provides the network with multiple routes to efficiently and effectively reorganize itself to detect, interpret, and react within its environment. The following will provide an overview of the biological mechanisms which steer the nature of local nonlinear interactions (culminating into complex global emergence).
2.1. Synaptic Plasticity
The following is a biological review of the various modes of connectivity and plasticity in the brain engendering nonlinear interactions. Although by no means exhaustive, our review represents a fundamental foundation with references that convey the necessary important takeaways, that is, the variety of biological mechanisms for connectivity and how they can change over time to support dynamic brain function. Synapses are not stationary over time. They are highly dynamic, entailing a variety of presynaptic and postsynaptic mechanisms capable of changing over time to fine tune the overall efficacy of synaptic transmission and corresponding synaptic strength [
80,
81]. Thus, synaptic plasticity confers the highest-resolution modus operandi in the brain for controlling and modulating interactions between constituents with the smallest temporal and spatial scales possible. Presynaptic plasticity includes modulation of presynaptic intracellular Ca
++ concentrations. This is primarily controlled by the function of voltage-gated calcium channels, which, when activated upon an incoming action potential, allow for the influx of Ca
++ inside the cellular presynaptic domain. Correspondingly, Ca
++ serves as a secondary messenger [
82]. As calcium has a high reactivity with a variety of substances, it serves as the ideal secondary messenger to relay information. Thus, biological form manipulates Ca
++ reactivity to engender binding affinity upon different calcium-binding proteins. In the presynaptic cell, calcium forms a large signaling complex with SNAREs and associated proteins, triggering the binding of synaptic vesicles (containing neurotransmitters) with the membrane and consequent release of neurotransmitters within the vesicles [
83]. Thus, regulation of voltage-gated calcium channels in the presynaptic domain has a significant influence on synaptic strength [
84]. Furthermore, residual Ca
++ from prior activity can influence vesicle release [
85]. The quantal release of neurotransmitters freely diffuses across the synaptic space. Diffusion of neurotransmitters implies that they stochastically bind upon receptors in the postsynaptic domain. Probability of neurotransmitter binding is dependent on total amount or concentration of neurotransmitters [
86]. Larger amounts of released neurotransmitters result in a higher concentration of neurotransmitters in the synaptic space, corresponding to an increase in the probability of greater numbers of activated receptors, resulting in an interaction with greater magnitude between pre and postsynaptic cells. Therefore, factors such as Ca
++ concentration modulate synaptic strength by influencing vesicle release and, correspondingly, the total quantal number of released neurotransmitters. Furthermore, within the presynaptic domain, a pool of readily releasable vesicles is maintained to, as the name suggests, be released at a moment’s notice upon action potential arrival (triggering Ca
++ influx and consequent release of vesicles) to pervade the synaptic cleft with neurotransmitters. If these stores are exhausted by repetitive, higher-than-normal action potential firing, this may result in an overall decrease in the number of vesicles released, consequently reducing the concentration of neurotransmitters and vice-versa; factors that replenish or sustain a larger pool of readily releasable vesicles can increase the concentration of neurotransmitters [
87]. Extrapolating from this, synaptic strength can be influenced by factors that control the concentration of neurotransmitters in the synaptic cleft. Thus, enzymatic machinery responsible for reducing the neurotransmitter concentration in the synaptic cleft to reduce the neurotransmitter activation time also influences the time course of synaptic strength [
88]. This is an essential mechanism to terminate a signal, thereby offering additional degrees of flexibility in fine tuning synaptic dynamics.
Furthermore, there are multiple neurotransmitter reuptake mechanisms (or neurotransporters) responsible for removing neurotransmitters in the synaptic cleft [
89]. These can also be utilized for future neurotransmitter release; thus, while influencing the concentration of neurotransmitters in the synaptic cleft, they can also alter the storage of readily releasable vesicles, consequently influencing the possible concentrations of neurotransmitters in the future. Reuptake can be undertaken by neurons and glia cells alike and is driven by neurotransporters, which can offer additional degrees of freedom to modulate synaptic connection strength by altering neurotransmitter concentrations [
90,
91]. Additionally, it must be recognized that non-neuronal glia cells (such as astrocytes [
92]) can also modulate synaptic transmission [
93,
94]. Their importance, along with that of other types of glial cells, such as astrocytes, oligodendrocytes, and microglia, has recently come to light, and as research progresses, this further illuminates the importance of a variety of cells (having clear dynamical roles) previously considered to have relatively stationary roles in the dynamical ensemble of a neural network [
95,
96,
97].
Synaptic strength modulation by postsynaptic mechanisms is accomplished by controlling the availability and number of receptors on the synaptic site. A greater number of available receptors results in a higher probability that freely diffused neurotransmitters (1) bind upon receptors and (2) elicit a post synaptic response. In other words, receptor amount and availability are directly correlated with synaptic strength. Therefore, postsynaptic plasticity mechanisms operate by modulating the properties of postsynaptic receptors. Receptor subtypes such as AMPAr and NMDAr play significant, dynamical roles in controlling factors such as receptor expression and availability [
98]. Intracellular Ca
++ concentrations once again play a large role as secondary messengers in modulating the expression of receptors. CaMKII and calcineurin are two examples of calcium-binding proteins, where the former typically initiates phosphorylation, typically resulting in long-term potentiation (synaptic strengthening), whereas the latter initiates dephosphorylation events that often lead to long-term depression (weaking of synapses) [
99,
100]. Of utmost relevance to synaptic plasticity, the intracellular Ca
++ concentration regulates the expression of receptors. A higher Ca
++ concentration increases the probability of Ca
++ binding and activating protein units, resulting in AMPAr exocytosis [
101]. A larger number of AMPAr results in a greater cumulative cross-sectional available area of receptors. Ergo, the flux of ions across the membrane multiplied by the cumulative greater cross-sectional area of the receptors (due to AMPAr exocytosis) results in an overall larger increase in postsynaptic potential, that is, a greater level of influence between neuron cells through a stronger degree of coupling [
102].
NMDAr Mg
++ blockage and relief of blockage via membrane potential excitation are at the core of controlling the direction and magnitude of plasticity [
103]. This is based on temporal correlation of presynaptic and postsynaptic neuron firings [
104]. Thus, the timing of interactions between presynaptic and postsynaptic neurons determines the overall amount of available NMDAr (relieved of Mg
++ blockage). This is reflected by Hebbian learning rules illustrated through spike-timing-dependent plasticity (STDP). The general takeaway is that neurons that fire together wire together by increasing their mutual coupling strength [
105]. The subtlety of this phenomenon has been pruned over time, and whereas the popularization of STDP clarifies how temporal correlation of pre- and postsynaptic firing coincidence directs synaptic strength, it must be understood that this is a simplification of the actual underlying molecular and cellular mechanisms [
106,
107]. Although this simplification can be a helpful analogy, neglecting the fundamental details obscures the full repertoire of nonlinear dynamics supplanted by synaptic mechanisms. Imprecise truncation of the local nonlinear interactions renders severe alterations in global form and function, as opposed to more comprehensive incorporation of the full repertoire of nonlinear local interactions.
When a postsynaptic cell fires after the presynaptic cell, there are greater numbers of unblocked NMDAr on the postsynaptic site that increase the overall receptor cross-sectional area for this uniquely Ca
++-permeable receptor. Therefore, if presynaptic neuron firing releases neurotransmitters that diffuse across the synaptic site at the time when NMDAr are unblocked, ligand activation of the NMDAr results in an increased level of Ca
++ influx. Consequently, intracellular Ca
++ levels rise, increasing the probability of Ca
++ secondary messengers activating AMPAr exocytosis. In some situations, different subtypes of AMPAr increase on the membrane that are also permeable to Ca
++, thereby increasing the probability of elevated Ca
++ levels [
108]. Furthermore, intracellular Ca
++ concentrations can be modulated by internal release of calcium from intracellular stores. These can be controlled by metabotropic receptor activation [
109]. Additionally, multiple types of receptors are expressed, offering a variety of mechanisms across a range of time scales. Of these, ionotropic and metabotropic receptors [
110] are some of the most prevalent and widely studied. Ionotropic receptors (or ligand-gated ion channels) typically operate on a shorter time scale, whereas metabotropic (or G-protein-coupled receptors) have longer activation times and work over a longer time-period due to the additional metabolic steps necessary in between agonist binding and elicited postsynaptic response via ion flux. The variety of receptors operating on different time scales further engenders nonlinear interactions amongst constituents. There is a wide multitude of forms of synaptic plasticity used in a variety of brain regions. The objective of this paper is not to provide a comprehensive description of all these forms but simply to provide the general foundations for the various modes of synaptic plasticity in the brain; references [
111,
112,
113,
114] provide more comprehensive reviews of synaptic plasticity.
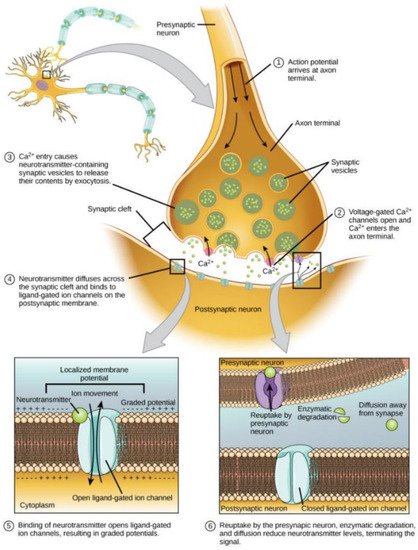
Figure 1 [
115] displays a synaptic diagram with specifications of its relevant components to illustrate how, ultimately, neurotransmitter concentration and cumulative numbers of available receptors influence the magnitude of interactions between neurons in terms of regulating total ionic flux.
Figure 1. A simplified illustration of the synapse. Magnitude of interaction is determined by concentration of neurotransmitters and cumulative availability of receptors. Direction of interactions (excitatory or inhibitory) is typically controlled by the type of neurotransmitter released. Thus, factors that influence these parameters control synaptic strength. As synapses are housed on axonal and dendritic structures, their properties also have significant influence on synaptic strength. Reproduced with permission [
115].
2.2. Axonal and Dendritic Structural Plasticity
Axonal and dendritic physiology further provide additional degrees of freedom to modulate connections between neural agents via structural plasticity [
116,
117]. For example, synapses are housed on dendritic spines, which offer stability to the synapse while supplying it with essential resources to support its activity. Thus, dendritic spine growth must follow synaptic dynamics. Should a synapse be particularly active, dendritic spine growth must increase to support a power-hungry synapse and vice-versa [
118]. Dendritic spines provide structural support to synapses and can supply necessary resources which help in facilitate dynamical receptor functions (e.g., modulating receptor expression). Furthermore, dendritic spines help transmit electrical signals to the neuron’s cell body, helping process input further. On the presynaptic end, axonal boutons also support presynaptic sites to supply synapses with resources, such as neurotransmitters, thus supporting synaptic strength [
119]. Furthermore, dendritic branching [
120] offers additional degrees of computation to neurons, increasing the degree of freedom with which neural connectivity can maneuver. Axons confer additional methods for plasticity on a larger scale [
121,
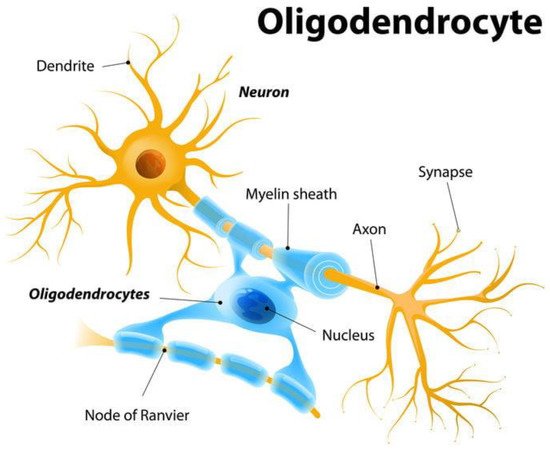
122]. The axon is responsible for transmitting an action potential from cell body to axon terminal at its presynaptic sites. Myelin sheaths, produced by oligodendrocytes, are insulating layers encompassing axons made of protein and fatty substances that coat the axon to speed up action potential transmission through saltatory conduction (see
Figure 2 [
123]). Naturally, the distribution of myelin carries significant implications for the temporal evolution of signal transmission throughout the brain. Axonal arborization can be particularly extensive, connecting a variety of brain regions. Hence, manipulating the signal transmission speed along axonal white matter tracts by controlling the distribution of myelin confers the ability to drastically change firing pattern interactions between relatively larger-scale (with reference to synaptic mechanisms) brain regions [
124]. This from of plasticity is highly prevalent to adaptation in the adult brain [
125]. Adaptive myelin plasticity modulates the growth and formation of myelin along axon bundles throughout cortical regions to modulate the speed and efficacy of information transfer. In other words, this can change the character of spatiotemporal frequencies of brain activity. High-resolution synaptic connections have been pruned through earlier experiences, restricting how flexible conformation changes can occur at this level. However, adaptive myelination is a form the adult brain commonly uses to refine signal transmission, albeit at a lower spatiotemporal resolution. This explains how young children, with fresh synapses, can learn new concepts to such a high level of resolution. Adults are still capable of learning through adaptive myelination; however, due to synaptic pruning in their youth, the resolution of detail that they can learn is not as refined. For example, an adult can learn a new language; however, it will be far more difficult to learn and achieve the subtleties of a native language speaker’s accent.
Figure 2. Myelin sheath distribution by oligodendrocytes on axons to speed up action potential conduction. Adaptive myelination, by controlling distribution of myelin, confers larger-scale modulation of signal transmission dynamics, as opposed to synaptic plasticity mechanisms. Activity-dependent control of myelin distribution along white matter tracts of the brain (connecting different regions) temporally modulates signal transmission, resulting in reconfiguration of connections between larger-scale brain regions. Reproduced from Shutterstock [
123].
The direction of such interactions is typically determined by the type of neurotransmitter used. For example, glutamate is used in excitatory neurotransmission, whereas GABA is used in inhibitory interactions. Furthermore, neurotransmitters can elicit modulatory responses. These can entail a combination of excitatory and inhibitory action [
124,
125,
126,
127] by being able to release multiple neurotransmitter types.
It must be noted that the preceding mechanisms are only the tip of the iceberg, providing a fundamental foundation to describe the various levels of intricate, detailed manipulation in neural connections fueling the emergence of complex brain dynamics. For a more comprehensive review where this subject matter is the main focus, the literature referenced above is recommended. In the context of this paper, it is important to recognize that the variety of biological connectivity entails a wide range of capabilities in precisely fine tuning the nature of nonlinear dynamic interactions across the dynamical hierarchy of the brain.
Furthermore, previous studies have established a preliminary qualitative understanding regarding the underlying biological machinery of the brain. However, to develop further refined insights, these qualitative biological interactions must be quantitatively expressed to precisely encapsulate the inherent nonlinearity and coupling. This can enable further progress by addressing current limitations. For example, current methods lack the resolution and quantitative precision of enumerating global brain dynamics. A theoretical, numerical model describing coupling at the level of synapses can aid in providing a more precise quantitative description. As these global properties are a result of the nonlinear couplings between constituents, defining the degree of coupling can aid in producing refined models and, consequently, a deeper understanding of the brain.
2.3. Quantifying Dynamical Local Coupling
Coupling strength, or interaction magnitude, at the synapse is determined by a combination of highly nonlinear processes, such as (1) the concentration of neurotransmitters in the synaptic cleft and (2) the total number and availability of receptors on the postsynaptic site. Neurotransmitter binding upon receptors is not deterministic but inherently stochastic. Therefore, the concentration of neurotransmitters in the synaptic cleft and the total number of available receptor binding points on the postsynaptic membrane can be used to generate a probability of receptor activation. The probability of receptor activation can be expressed in terms of the total cross-sectional area of receptors that allow for the influx of ions. Using fundamental diffusion principles formulated through Fick’s laws, the flux of ions can be quantified with regard to the established electrochemical gradient between the intra- and extracellular space. Thus, the flux of ions multiplied by the total cross-sectional area of receptors corresponds to the total amount of ion influx across the membrane. Incorporating this value with the electrochemical gradient, temporal iteration time and charge for corresponding ion species summed over all synaptic points can represent the voltage fluctuations of a neuron over time. Equation (1) provides a preliminary governing dynamical equation to quantify coupling in terms of postsynaptic potentials. This can serve as foundational coupling law to determine whether a neuron will fire or not based on its synaptic inputs. Voltage (
Vi), the energy per unit charge at the next time step, is equal to the voltage at the previous time step plus the summed (over all synapses and ion species respectively) product of the electrochemical gradient (
∇μ) in joules per mol; the total cross-sectional area of the open ligand-gated channel (
α); the flux of ions per area per unit time,
Jflux; and the charge per ion species,
qion. This coupling law defines the dynamical voltage fluctuations of a neuron with reference to its synaptic inputs.
It must be recognized that the electrochemical gradient and flux due to diffusion are relatively stationary. Hence, the term that represents the dynamical nature of coupling is α. This term is fundamentally nonlinear, as it is equal to the total cross-sectional area of the open ligand-gated channels, which is simultaneously dependent on pre- and postsynaptic mechanisms, such as the concentration of neurotransmitters which probabilistically bind upon postsynaptic receptors that may or may not have a voltage-dependent Mg++ blockage. Hence, as a product of the variety of plasticity mechanisms, α is stochastic and highly nonlinear. It can influence (1) the concentration of neurotransmitters and (2) the number and availability of receptors on the postsynaptic site. It must be noted that this equation is a foundational factor in quantifying coupling in the brain, particularly on the micro scale. Additional coupling terms, such as adaptive myelination, must be incorporated to comprehensively account for coupling on a larger scale. Furthermore, additional revisions are required to explicitly incorporate and quantify the various biological mechanisms that modulate the dynamical trajectories of neural postsynaptic potentials. Regardless, quantifying coupling at the microscopic scale is a necessary first step towards a more complete model. Nonetheless, the underlying philosophy of this equation can be utilized to aid in quantifying complex local voltage fluctuations due to interactions amongst neuronal constituents.
2.4. Local Interaction-Induced Global Characteristics
As described, there is a broad distribution of plasticity mechanisms influencing functional, structural, temporal, and spatial behavior of neural interactions from the micro to macro scale. Furthermore, these mechanisms are not implemented in isolation but incorporated simultaneously, enabling different degrees of maneuverability in connection strength and direction. Consequently, these local interactions are highly nonlinear [
128]. When combining these various components, global network dynamics are consequently nonlinear and, when undergoing complex dynamical evolutions, can display chaotic characteristics [
129]. These are necessary for fluid multivariable adaptation, as the environment consists of a variety of nonstationary conditions requiring complex physiological form to not only ensure survival but to optimize conditions of survival (e.g., subcortical motivations, steering the quality of life, and gauging reproduction thresholds). Evolutionary adaptation has encoded a fundamental configuration for neural connectivity within the brain, resulting in its natural hierarchical order from birth. Life experiences over time fine tune neural connectivity with adaptive plasticity mechanisms to mold the adult brain. This refines a neural network’s instinctive response to environmental stimuli in attempts to optimize its survival.
From a higher-level perspective, global brain dynamics are the processes that steer the network to adapt within the constraints of nature. These are not static in time but highly time-variant from the micro to macro scale, structured in intricate layers of modular connectivity, allowing for coordinated, efficient, dynamic organization [
130,
131,
132,
133,
134]. Therefore, unique microstate configurations (the exact individual behaviors of network constituents and the degree coupling between these network nodes produced by physiological configurations) determine the global macrostate emergent forms. Thus, the brain is a highly adaptive network whose characteristics change over time to interact with a nonstationary environment. Adaptation entails changing the global properties of a network system over time in response to varying external input posed by environmental conditions. These macroscopic dynamics exhibit transitions from distinct states of global brain function to ensure stability (i.e., survival) in accordance with external situations. Different environmental scenarios, such as scavenging for resources, such as food and water; reading social communication cues; fight or flight response towards predators; sleep; and abstract conceptual thinking, necessitate a variety of distinct global brain functions created by respective microstate configurations of cumulative local neuron interactions [
135]. As previously mentioned, the variety of macroscopic distributions (global brain states) is the result of the microscopic configurations of the ensemble’s constituents, i.e., the cumulative behaviors and interactions between neurons mediated through their connections with one another, which regulate neural dynamical activity. Therefore, brain macrostate transitions in the form of adaptations to new environmental stimuli are also facilitated by changing the respective microstate configurations. In other words, this corresponds to changing the biological mechanisms between neurons and glia cells by changing the expression or availability of receptors between neurons or adjusting the concentration of neurotransmitters in the synapse [
136]. This is similar to how global phase transitions are facilitated by a change in the interactions between molecular constituents [
137]. Brain network state transitions are directed by modulating the strength of synaptic and structural couplings between neurons, steering the magnitude and direction of local neuronal interactions that culminate into emergent dynamical trajectories [
138]. The governing philosophy of a brain network is that the global level forms and their changes over time are the result of the local-level dynamical interactions of the constituents that compose the ensemble. Hence, the particular microstate configurations in terms of the exact myelin distribution across white matter fiber tracts, dendritic branching, and spine characteristics, along with synaptic efficacy determined by the product of neurotransmitter concentration and receptor availability, cumulatively engender highly nonlinear connectivity. These relationships between network constituents are highly nonlinear and recursively couple upon one another across the temporal and spectral scales of brain activity capable of producing chaotic characteristics.