In breast, nasopharynx, gastric, pancreatic, pulmonary, renal, lung, ovarian, and thyroid cancers, lysyl oxidase and collagen were found to influence the architecture of the ECM, creating a favorable microenvironment for tumor development and progression [
26,
27]. Leeming et al., 2019 [
28] found that healthy humans presented serum LOXL2 enzyme levels of ≈46.8 ng/mL. In patients with breast, colorectal, lung, ovarian, and pancreatic cancer, the levels of LOXL2 in serum were significantly elevated, varying between 49 ng/mL and 84 ng/mL. Regarding patients with breast cancer, serum LOXL2 levels were elevated by 218% compared to healthy controls [
28]. A study from Janyasupab et al. (2016) [
29] measured LOXL2 levels in human serum, plasma, and urine. The researchers found differential LOXL2 concentrations in patients with breast cancer (≈ 2.7 µM in blood; ≈ 40 µM in urine), when compared to the cancer-free individuals (≈ 0.6 µM in blood; ≈ 25 µM in urine).
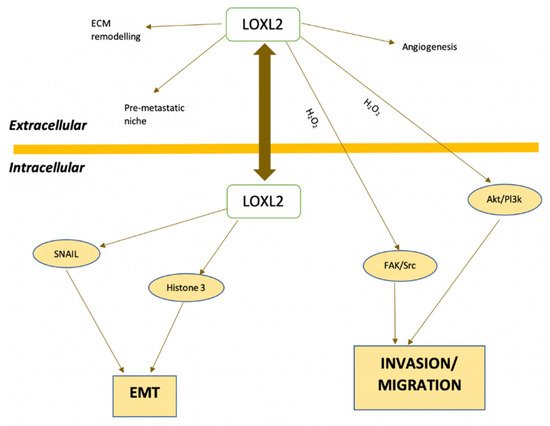
Normal breast tissue has lower levels of LOXL2 expression and this protein is found in the stroma and luminal layer of epithelial cells. Conversely, breast cancer tissues show increased LOXL2 expression, located intracellularly in the cytoplasm and cell nuclei, and extracellularly in the ECM [
39,
51]. Immunohistochemical studies demonstrated that approximately 60% of basal breast carcinomas have increased intracellular LOXL2 with perinuclear distribution, associated with mRNA overexpression [
39]. In addition, there is an increase in LOX expression in metastatic tissues compared with primary tumors [
31]. Using immunohistochemistry, Ahn et al. (2013) [
50] demonstrated that LOXL2 is an independent prognostic marker of metastatic disease and death in patients with breast cancer. In addition, LOXL2 is an independent prognostic factor for overall survival (OS) and metastasis-free survival (MFS) in breast cancer (hazard ratio of 2.27 and 2.10, respectively) [
50].
Considering the therapeutic potential of blocking LOXL2 in cancer treatment, several inhibitors of this protein have been developed. The next sections summarize the state of the art of the discovery of LOXL2 inhibitors.
4. LOX Inhibitors
4.1. β-Aminopropionitrile (BAPN)
As mentioned in
Section 2, BAPN was the first LOX inhibitor to be identified. BAPN inhibits intramolecular and intermolecular covalent cross-linking of collagen and elastin connective tissue proteins [
6]. In fact, BAPN is described as a potent and irreversible non-specific LOX inhibitor, which also has an affinity for other amine oxidases [
54].
LOX inhibitory activity of BAPN was evaluated in an assay developed in rats [
55]. The authors found that doses of BAPN ranging from 1 to 40 mg per 100 g bw were efficient in inhibiting LOX activity from 6 and up to 48 h. They also studied the kinetics of both BAPN and its major metabolite, the cyanoacetic acid (CAA), in the same model. After a single intraperitoneal dose, most of the BAPN was excreted unchanged in the urine, while the rest was metabolized slowly in CAA [
55].
Tang et al. (1983) [
56] suggested a possible mechanism for the interaction between BAPN and LOX. By using BAPN with isotopically labeled carbons, the authors found that this molecule covalently binds to LOX to equivalent extents and in parallel with the development of inactivation, without the elimination of nitrile moiety. The copper of the enzyme is not altered upon interaction with BAPN, and BAPN is not processed to a free aldehyde product. The suggested inhibition mechanism involves the formation of a covalent bond between an enzyme nucleophile and a ketenimine formed from BAPN.
This LOX inhibitor has shown anticancer properties in several in vitro and in vivo models of different cancer types. For example, Yang et al. (2013) [
57] demonstrated that BAPN (500 µM) blocked the hypoxia-induced invasion and migration capabilities of cervical cancer cells. Zhao et al. (2019) [
58] have shown that the inhibition of LOX by BAPN in BGC-823 gastric cancer cells inhibits the expression and activity of matrix metalloproteinases 2 and 9.
Regarding the effects of BAPN in breast cancer, Cohen et al. (1979) [
59] used rats with breast tumors induced by 7,12-dimethylbenzanthracene. The authors found that BAPN inhibited the collagen cross-link and promoted an 82% decrease in tumor formation and a significant reduction in tumor volume. In another experiment, luciferase-expressing breast cancer cells (MDA-MB-231-Luc2) were injected in mice to explore the effects of BAPN in invasion to other organs [
60]. The results show that BAPN reduced the appearance of metastases. The number of metastases was decreased by 44%, and 27%, when BAPN treatment was initiated the day before or on the same day as the intra-cardiac injection of cancer cells, respectively. However, BAPN showed no effect on the growth of established metastases. The authors concluded that LOX inhibition might be a useful strategy for metastasis prevention [
60].
Another potential use of BAPN and LOX inhibitors was suggested by Rachman-Tzemah et al. (2017). Increased LOX activity and expression, fibrillary collagen cross-linking, and focal adhesion signaling observed after breast tumor resection contribute to increasing the risk of lung metastases [
61]. LOX pharmacological inhibition using BAPN or an anti-LOX antibody prior to surgical intervention was able to reduce lung metastasis after surgery and increased animal survival in a murine model of breast cancer [
61].
Despite the interesting results obtained with BAPN, this molecule lacks suitable sites for chemical modifications [
62]. This fact does not facilitate the preclinical optimization. Conversely, novel classes of LOX enzyme inhibitors do not present this drawback, making it an advantage in drug discovery, as described in the subsequent sections.
4.2. Copper Chelators
Cox, Gartland, and Erler (2016) [
63] proposed an indirect approach for LOX inhibition, using tetrathiomolybdate (TM). This is a potent copper chelator that targets the catalytic activity of LOX by binding to copper and depleting it. Copper has a significant influence on the functional activity of LOX, although it does not directly interfere with its expression levels. In preclinical studies, tetrathiomolybdate showed antiangiogenic activity, antifibrogenic and anti-inflammatory actions. A recent study found that copper was elevated in fibrotic kidney tissue and such increase promoted LOX activity and extracellular collagen cross-linking [
64]. Copper chelation by TM leads to a decrease in activated LOX protein [
64].
Another copper chelator initially suggested to be a LOXL2 inhibitor is D-penicillamine (D-pen) [
65]. Contrarily to BAPN, D-pen structure has a secondary amine [
32]. D-pen drastically inhibits rhLOXL2 activity at a concentration of 10 µM [
52]. However, despite some conflicting data regarding its mechanism of action, as a copper chelator, D-pen is considered a non-selective inhibitor of LOXL2 enzyme (reviewed in [
32,
65]). In an orthotopic breast cancer mouse model, D-pen showed no effect on tumor growth rate. However, mice bearing tumors treated with D-pen displayed fewer lung and liver metastases than untreated mice [
52]. Accordingly, in a transgenic breast cancer model, D-pen treatment led to a decreased development of lung metastases when compared to control mice [
52].
Despite some encouraging results obtained with TM and D-pen, the chelation of cooper is not selective for LOXL2 or even for enzymes of the LOX family. Since copper ions play a part in several biological processes and are implicated in different enzymatic reactions [
66], the use of such chelators will likely disturb other biological functions.
4.3. LOX/LOXL2 Selective Inhibitors
Following the discovery of BAPN, some compounds have been developed with LOX and LOXL2 inhibitory activity and favorable pharmacokinetics parameters. However, targeting LOXs with specific small molecule inhibitors presents a challenge due to the lack of crystalline structures, since only the LOXL2 crystalline structure is available.
The LOXL2 inhibitors PXS-S1A and PXS-S2A are haloallylamine-based molecules (the structures are not disclosed) [
67]. PXS-S1A is a first-generation LOX inhibitor that exhibits an identical activity and selectivity when comparing to BAPN. The pIC
50 values against LOXL2 are 6.8 ± 0.2 for PXS-S1A and 6.4 ± 0.1 for BAPN, and the two compounds also have similar pIC
50 values when tested against the native human LOX enzyme. PXS-S1A allows for structural modifications that can be introduced to improve the inhibiting potency of LOX/LOXL2, thus leading to significant increases in selectivity. Chemical modifications of PXS-S1A led to the development of PXS-S2A, a potent and specific inhibitor of LOXL2 (pIC
50 = 8.3 M). The discovery of PXS-S2A established the basis for dissecting the functional role of LOXL2 in the progression of solid tumors such as breast cancer [
67]. These two LOX/LOXL2 inhibitors reduced the in vitro 2D and 3D proliferation of the breast cancer cell line MDA-MB-231 in a dose-dependent way [
67]. This cell line has a high level of LOXL2 expression. Although authors also describe a significant impairment in 2D and 3D cell motility, the assays were performed under similar conditions to those that lead to a reduction in cell proliferation. Thus, implying that the observed reduction in cell motility may also be partially due to the reduced cell proliferation of cells treated with the compounds. Importantly, the authors observed a clear reduction of in vivo orthotopic MDA-MB-231 primary tumor volume and tumor cell proliferation upon treatment with PXS-S1A and PXS-S2A [
67]. All the above-mentioned inhibitory effects were more pronounced for PXS-S1A when compared with PXS-S2A.
Another class of LOXL2 inhibitors is the patented collection of diazabicyclo[3.2.2]nonanes with a des-primary amine group. Compounds of this class were tested in a transgenic mouse breast cancer model and led to a reduction in the formation of lung metastases (reviewed in [
32]).
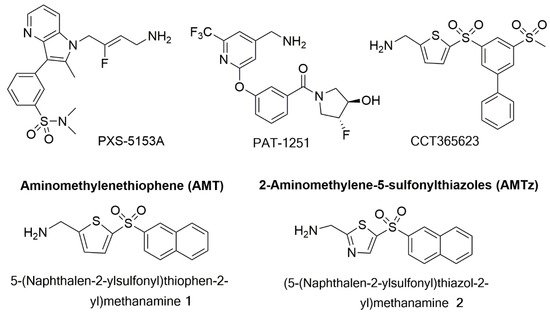
PAT-1251 (
Figure 5) was the first small molecule that acts as an irreversible LOXL2 inhibitor to advance to clinical trials (see
Section 7.5). This compound is a potent and highly selective oral LOXL2 inhibitor that is based on a benzylamine with 2-substituted pyridine-4-ylmethanamines [
65,
68].
Figure 5. Structures of PXS-5153A; PAT-1251; CCT365623; an aminomethylenethiophene (AMT), 5-(naphthalen-2-ylsulfonyl)thiophen-2-yl)methamine 1; and a 2-aminomethylene-5-sulponylthiazole (AMTz), (5-(naphthalen-2-ylsulfonyl)thiazol-2-yl)methanamine 2.
Epigallocatechin gallate (EGCG) is a trihydroxyphenolic compound that was suggested as a dual inhibitor of LOXL2 and transforming growth factor-β1 (TGFβ1) receptor kinase [
69,
70]. This compound induces the auto-oxidation of a LOXL2/3–specific lysine (K731) in a time-dependent manner that inhibits LOXL2 irreversibly [
69]. ECGC attenuates TGFβ1 signaling and collagen accumulation and was thus suggested as a possible therapeutic approach against fibrotic diseases [
69,
70].
Besides small molecules, the LOXL2 inhibiting strategies developed so far also include biological drugs. The anti-LOXL2 functional antibody named Simtuzumab is an IgG4 humanized monoclonal antibody, that is a non-competitive inhibitor of extracellular LOXL2 via allosteric inhibition by binding to the fourth SRCR domain. This antibody revealed beneficial effects in various preclinical models of fibrosis and cancer [
52,
71]. Moreover, Simtuzumab has been evaluated in several clinical trials, as detailed in
Section 7.5. While Simtuzumab targets noncatalytic regions of LOXL2, Grossman et al. (2016) [
72] developed specific antibodies that targeted the active site of this enzyme. Among those, the antibody clone designated GS341 displayed binding affinity to LOXL2 at the subnanomolar range and prevented the assembly of linear collagen fibers, producing visible variations in fibrillary collagen morphology. The GS341antibody was evaluated in a breast cancer xenograft model using MDA-MB-231 cells into immunocompromised SCID mice. Treatment with GS341 resulted in a decrease in tumor volume and in the number of lung metastases. In addition, the tumor fibrils in the GS341-treated animals were thinner when compared with the vehicle-treated ones [
72].