The use of biomarkers in cancer diagnosis, therapy, and prognosis has been highly effective over several decades. Studies of biomarkers in cancer patients pre- and post-treatment and during cancer progression have helped identify cancer stem cells (CSCs) and their related microenvironments. These analyses are critical for the therapeutic application of drugs and the efficient targeting and prevention of cancer progression, as well as the investigation of the mechanism of the cancer development. Biomarkers that characterize CSCs have thus been identified and correlated to diagnosis, therapy, and prognosis. However, CSCs demonstrate elevated levels of plasticity, which alters their functional phenotype and appearance by interacting with their microenvironments, in response to chemotherapy and radiotherapeutics. In turn, these changes induce different metabolic adaptations of CSCs.
1. Introduction
It is believed that uncontrolled progression of tumor cells is generated by a small population of cancer stem cells (CSCs) that possess the capability for self-renewal and pluripotent differentiation into multiple cancer cell types [
1]. CSCs are hypothesized to persist in cancers and cause metastasis, therapy resistance, and post-operative recurrence by producing new tumor cells. CSCs can survive many commonly employed treatments [
2]. Accordingly, targeting CSCs should provide new therapies to improve survival of cancer patients. Moreover, this research field may identify the heterogeneity of tumor cell populations and their genetic, epigenetic, and microenvironmental diversification.
The existence of CSCs was first identified in acute myeloid leukemia in 1997 [
3], although the terminology was first employed by Reya and colleagues in 2001 [
4]. CSCs were then found in glioblastoma [
5], breast carcinomas [
6,
7], gastric cancer [
8], and colorectal cancer [
9]. However, until now, the mechanisms that drive the intracellular dysregulation of CSCs to malignancy have remained unclear. Biomarkers of these CSCs are thus critical for investigating the mechanisms by which CSCs can develop into neoplasia interacting with the surrounding cells. In addition, these markers are useful for both identifying the heterogeneity of CSCs and determining their cell fates.
2. Perspective on the Therapeutic Use of Biomarkers
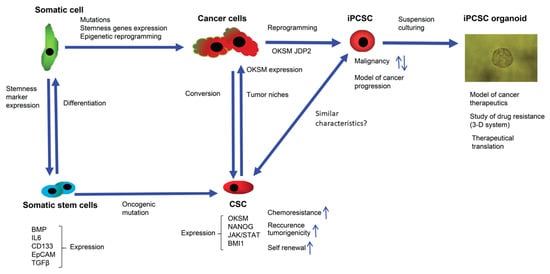
Based on existing data, the stream of the commitment of somatic cells and cancers cells to undergo reprogramming to produce normal stem cells and CSCs, as well as induced stem cells, can be summarized, to understand their stemness and their biomarkers (Figure 1).
Figure 1. Cancer initiation and the process of cell stemness induction (reprogramming) employs overlapping molecular signaling and epigenetic pathways. Expression of transcription factors—such as OCT4, KLF4, SOX2, NANOG, and JDP2—is necessary for cancer initiation, along with genetic mutations and epigenetic changes. In somatic stem cells, for example MSCs, an increased expression of CD133 by the JAK-STAT 3 pathway activates the migration of MSCs to cancer cells and can increase the number of CSCs. CSC niche factors that induce self-renewal of CSCs stimulate angiogenesis and recruit other cells producing additional factors related to tumor metastasis. Patient-derived iPCSC organoid models are useful for examining the mechanisms of drug resistance and cancer progression.
In this review, we have focused on reprogramming factors as putative universal biomarkers of stem cells, because they play a role in stem cell pluripotency and their interactions with various co-factors can enable transcriptional versatility during development [
95,
96]. CSCs are known to be a small population with self-renewal capacity and differentiation potential that confers tumor relapse, metastasis, heterogeneity, multidrug resistance, and radiation resistance [
97].
Stemness genes such as
OCT4,
SOX2, and
NANOG; some signaling-related stemness proteins, including WNT, NF-κB, NOTCH, HEDGEHOC, JAK-STAT, PI3K/AKT/mTOR, TGF/SMAD, and PPAR; as well as cell communication microenvironments such as vascular niches, hypoxia, TAMs, CAFs, ECMs, and exosomes are critical to the regulation of CSCs. Drugs, small molecules, vaccines, antibodies, and chimeric antigen receptor-T (CAR-T) cells targeting these pathways have been generated to target CSCs [
98]. How can stemness genes in normal cells be altered to cancer stemness genes using driver mutations? As previously described, the reprogramming factor, OCT4, is required for the generation of liver CSCs with the oncogene,
c-JUN [
23,
42]. Moreover, the AP-1 repressor, JDP2, plays a dual role in the reprogramming of cancer cells [
41]. Why do we observe the reversed role of JDP2? We found that the status of TP53 in cancer cells is critical for the generation of oncogenic and anti-oncogenic function [
1,
41,
42,
45]. This critical function of TP53 mutation in cancer cells was also reported to generate the reprogramming by other authors [
43,
44]. Expression of normal p53 in the original somatic cells is critical for normal reprogramming [
99,
100,
101]. Therefore, the status of TP53 and/or the methylation status of
p16Ink4a should be examined [
1,
44,
45]. Stemness genes are mutated or not during the cellular reprogramming of cancer cells to generate CSCs. Therefore, the role of at least the
OCT4,
SOX2, and
NONOG genes in the metabolic reprogramming, cell plasticity, and trans-differentiation processes should be clarified in future studies. Indeed, the functional role of stem cell factors in cancer commitment should be further investigated. Recently, the role of SOX2 has been reported [
102].
For example, the introduction of SOX2 in prostate cancer induces stem-like characteristics but uses different metabolic pathways and interacts with different target gene products. The oncogenic role of SOX2 is confirmed further by several studies exhibiting SOX2 dependent alteration of cell growth, invasion ability, and chemo-resistant activity beyond tumor types [
103,
104]. de Wet et al. demonstrated that most target genes of SOX2 regulation in prostate cancer are non-overlapping with targets of SOX2 in human ESCs, and identified different
cis elements within what appear to be similar target genes.
In addition, SOX2 employed the activities of canonical reprogramming, glycolytic, and oxidative phosphorylation reactions in prostate cancer cells different from those associated with canonical SOX2 function in ESCs. This suggests the relocation of SOX2 to novel oncogenic drivers during progression of prostate tumor [
102]. Thus, stem cell markers are not universal, and are context dependent. Some reports pertained to the role of SOX2 in cancer metabolism [
93]. Glucose metabolism in cancer exhibited a unique bioreaction whereby the main energy source of primary prostate cancer cells relied on oxidative phosphorylation and shifted toward a reliance on glycolysis for ATP generation during the metastatic stage of cancer. By contrast, the metabolic profile of pluripotent ESCs is featured by high glycolytic activity to compensate cell proliferation, in particular, during hypoxic state of the inner cell mass before implantation of the embryo [
105]. However, cancer cells use different metabolic alterations and nutrient adaptation during metastatic stages that lead to tumorigenesis. To date, there is little data supporting a mechanistic link between SOX2 and mitochondria and glucose metabolism directly [
106]. It is thus important to identify each master stem cell factor and its role during different embryonic and cancer stages. Accordingly, these studies indicate that reprogramming procedures in cancer cells mostly lead to the generation of malignant cancer-initiating cells that acquire stem cell-like properties. Furthermore, studies of some stem cell reprogramming factors such as SOX2 have revealed new mechanisms by which they enable metastatic progression, cell-lineage plasticity, and therapy resistance. The hallmark of CSCs can be a combination of different characteristics such as phenotypical and metabolic markers that displayed a kind of signature which can be used as new target. These new CSC gene axes and the stemness genes might be representative of the new CSCs markers.
The identification of cellular plasticity and biomarkers are useful approaches for the study of cancer stemness and new oncogenic pathway. Recently, Yang et al. [
107] developed a specific genetically encoded biosensor with 72 barcodes (or 128 biosensors) expressed in each cell. In combination with the use of fluorescence, it is now possible to precisely identify the nature of cellular plasticity and discover new cellular pathways, connections, and mechanisms.
Another approach that has been suggested to be an effective novel cancer therapy is exosome-based delivery of cancer-suppressor proteins, microRNA (miR), or targeted drugs [
108]. Exosomes are membrane-bound extracellular vesicles that are much smaller than cells and produced in most eukaryotic cells [
109,
110]. MicroRNA in exosomes affects protein production in the recipient cells [
111]. Because exosomes express markers of the cancer cells of origin, clinical applications such as for biomarkers and therapies are realistic.
For example, exosomes containing miR-21 produced an effective downregulation of the
PDCD4 and
RECK genes in glioma cell lines [
112]. Furthermore, patient-derived exosomes loaded with paclitaxel were superior to paclitaxel-loaded liposomes as a cancer immunotherapy in lung cancer cell lines [
113], and exosomes carrying inhibitors of the self-renewal, differentiation, and tumorigenesis-related genes (e.g., miRNAs or siRNAs for
TGFβ,
Wnt,
Hippo, etc.) of CSCs are possible therapies for cancer treatment [
108]. Taken together, these data suggest that clinical application of exosome-based therapies for various cancers should be addressed in the near future.
3. Elusive Problems Faced to Eliminate CSCs
In efforts to remove CSCs effectively, a series of existing problems need to be addressed. (i) As mentioned, because the cancer signaling of CSCs is not specific and shares some pathways with normal stem cells, not all the regulatory factors with or without mutations that contribute to CSCs are appropriate for use as therapeutic targets. (ii) The precise features of many CSCs in specific types of cancers are not well characterized [
114]. (iii) Since the tumorigenesis activities of CSCs are determined in immune-deficient mice or animals in the absence of an adaptive immune system, they do not recapitulate the biological complexity of cancers in the clinic [
115]. (iv) Information about stem cell niches and the microenvironment are not sufficient. Studies of linkages between CSCs and microenvironments should be combined [
51]. (v) Novel studies of the signaling and regulatory mechanisms of cancer metabolism, epigenetics, and mitochondrial functions should be explored [
116]. (vi) Inhibitors, small molecules, and antibodies, as well as CSC-directed immunotherapy, need to be developed [
117]. These problems should be addressed to promote the promising application of biomarkers for CSC niches for cancer therapy.
4. Conclusions
Characterization of biomarkers in stem cells is critical for the identification of the biochemical and genetic events that trigger cancer development (Figure 1). Thus, for each biomarker, such as metabolic reprogramming factors, the links with specific cancer commitment and the associated signaling should be clarified to better understand the conversion of normal stem cells to CSCs. Furthermore, these biomarkers will be useful in the future application of therapeutic tools to treat human cancers. In the future, the identification of biomarkers for stem cell markers and the impact of alterations in signaling on these biomarkers during cancer commitment will be required to reveal the cellular mechanisms and key events in cancer induction that might be potential pharmacological targets.
This entry is adapted from the peer-reviewed paper 10.3390/jpm12050715