To model the in vivo developmental processes, 3D cell cultures were developed and categorized into different subtypes such as embryoid body (EB), gastruloid, spheroid, and organoid. EBs are aggregated differentiating embryonic stem cells (ESCs) or iPSCs. The cells in EBs are usually at early differentiation stages and do not organize into specific spatial patterns. In the cardiac stem cell field, EBs are used to generate lineage-specific cell types such as cardiomyocytes [
1]. Gastruloids consist of three germ layers and are used to model the embryo gastrulation process. In theory, gastruloids can develop into an entire embryo if an appropriate differentiation environment is provided; in a recent study, a differentiation condition was designed to enable the cultured mouse embryos to develop to the hindlimb formation stage [
2,
3,
4]. The spheroid is an aggregate of differentiated cell types or cancer cells. These cells do not self-assemble into the anatomical patterns observed in vivo and can be used to study the interactions among different cell types. Lastly, the organoid is a complicated cell aggregate in which the differentiated cells mostly belong to a specific organ and self-assemble into the in vivo-like anatomic patterns. Tissue-specific organoids, such as the cerebrum, liver, lung, and kidney, can be made with iPSCs or progenitor cells [
5,
6]. They usually have similar morphology and cell-type interactions to the in vivo organs, making organoids advantageous over other 3D models [
7]. However, as the emergence of multi-lineage organoids and certain organoids can only recapitulate part of the organ-like features, the distinctions between the different types of 3D cell cultures are becoming less clear.
Multiple hiPSC-derived heart organoid systems were recently reported and claimed to have developed in vivo-like heart features. The heart is the first organ to develop in humans, and its defects affect about 1% of births [
8]. Heart development originates from mesodermal cells, which later develop into cardiac progenitors in the two heart fields. First heart field (FHF) cells develop into the primitive heart tube, and second heart field (SHF) progenitors contribute to heart-tube elongation from both poles [
9,
10]. Meanwhile, the heart tube undergoes rightward looping, giving rise to the four-chambered heart. The heart chamber wall consists of three tissue layers: epicardium, myocardium, and endocardium. The heart also has other structures besides the chambers, including the inflow tract (IFT), the atrial ventricular canal (AVC), and the outflow tract (OFT) at the early stages, which contribute to the development of the chambers, valves, septum, and large vessels at later stages. The normal development of these structures is essential for generating a functional heart; if this process goes awry, it can cause congenital heart defects (CHDs) [
10].
2. Heart fields Formation
During mouse heart development, the two heart fields are specified sequentially and express marker genes such as
Hcn4 (FHF),
Isl1, and
Tbx1 (SHF) [
12,
13]. The FHF originates from the lateral plate mesoderm and mainly contributes to linear heart-tube formation. FHF formation is induced by signals from the adjacent ectoderm, endoderm, embryonic midline, and posterior regions. Bone morphogenetic protein (BMP), Fibroblast growth factor (FGF), Transforming growth factor β (TGFβ), and WNT signaling pathways were reported to be involved in this process [
14,
15]. The SHF is derived from the pharyngeal mesoderm and contributes to heart development from both inflow and outflow poles after the linear heart-tube stage. Additionally, the SHF is specified by signals from the surrounding pharyngeal endoderm and neural crest cells. While SHF progenitor cell survival, proliferation, and deployment were found to be dependent upon WNT, BMP, Hedgehog (HH), and FGF signaling [
15,
16], the posterior limit of SHF was reported to be determined by retinoic acid (RA) [
17]. Furthermore, TGF-β, HH, and FGF were reported to be important in zebrafish SHF progenitor cell proliferation and differentiation [
18] (
Figure 1A).
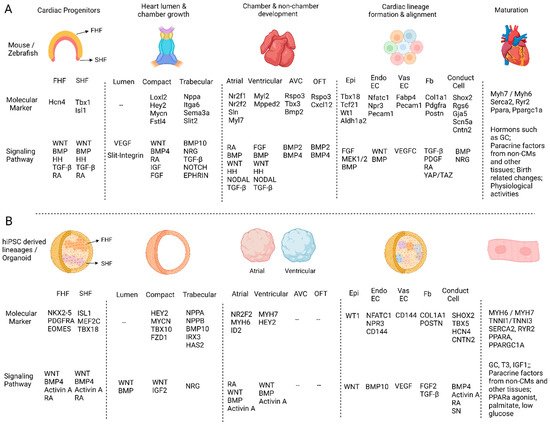
Figure 1. The comparison of molecular markers and regulatory signaling pathways in in vivo heart and human heart organoid development. (A) The major events during in vivo heart development and their molecular markers and regulatory signaling pathways. (B) The molecular markers and signaling pathways that have been reported to be important in regulating human iPSC-derived cardiac cell or cardiac organoid differentiations. “--” means non-applicable or no related studies have been reported.
The FHF and SHF were also discovered to develop in mouse 3D culture systems. Andersen et al. verified FHF/SHF-like cells in their mouse iPSC-derived organoids by showing similarities with embryonic FHF/SHF cells in their gene expression and differentiation potentials. They also found that the Bmp/Smad pathway and the Smad-independent BMP/WNT pathway specified FHF and SHF progenitors [
12], respectively. Recently, Rossi et al. described mouse embryonic stem cells (mESCs)-derived gastruloids that were found to have cells expressing the FHF/SHF markers and had a spatial distribution like the in vivo progenitors [
19]. Additionally, Lewis-Israeli et al. generated heart organoids from hiPSCs presenting characteristics of both heart fields in the same organoid. They identified
NKX2-5, PDGFRA, and
EOMES expression in the FHF progenitors and
ISL1,
MEF2C, and
TBX18 expression in SHF cells (
Figure 1B). These progenitors were described to respectively develop into left and right ventricular CMs based on the expression of
HAND1 and
HAND2 [
20].
Next, it will be interesting to analyze the detailed structures in the organoid heart fields, as the FHF and SHF in mouse embryos are specified into small segments through the differential expression of HOX genes and RA signaling, and each segment respectively develops into related heart anatomical structures such as atrial and ventricular chambers [
21]. Furthermore, these organoid heart fields do not seem to differentiate sequentially or form the specific spatial patterns seen in the heart fields in embryos, whose SHF locates dorsal and medial to the FHF to progressively contribute cells to both poles of the linear heart tube [
10]. With further detailed analysis and appropriate manipulations, we expect the organoid heart fields to develop in a similar temporal and spatial manner as seen in embryonic heart field development.
3. Heart Lumen Development
Through genetic screening in Drosophila, the heart tube lumen formation was found to be regulated by a Slit-Integrin signaling pathway, which regulates actin cytoskeleton alignment to promote cardiac cell polarization in lumen development [
22,
23]. In mice, live imaging analysis revealed that the heart lumen developed from a split of two endocardial endothelial cell (EndoEC) layers at the cardiac crescent stage. It also found that cell rounding was unlikely to initiate lumen formation as the cardiac crescent cells are still columnar when the lumen begins to develop [
24]. As part of the heart lumen developmental process, aorta lumen propagation initiates between stages 1S and 3S (E8.0), developing from adjacent endothelial cell (EC) contact after EC shape has changed. This process is regulated by VE-Cadherin and VEGF-A [
25]. In humans, the heart lumen develops from the fusion of two endocardial tubes, each of which has a hollow lumen derived from the cardiogenic cords [
26].
A recent study reported the induction of heart chamber-like structures in human heart organoids and found that Wnt-BMP signaling and transcription factor
HAND1 were both critical in this process [
27]. Further time-course analysis of organoid formation found that the lumen appeared after 2.5–3.5 days of differentiation at the cardiac mesoderm stage, which is earlier than when the mouse heart lumen develops in the cardiac crescent stage. Additionally, the study found that low WNT and Activin A levels can induce chamber formation with a partial inner lining of EndoECs. However, lumen formation does not seem to rely on the EndoECs, as the lumen can still form when the EndoECs developed on the outer organoid surface after VEGF treatment [
27]. Lewis-Israeli et al. and our study also generated organoids with chamber-like structures, and these chambers also developed independently from the EndoECs [
20,
28]. As chamber formation in the current organoids does not go through the same process as in vivo heart lumen development, there is limited value in modeling human heart lumen formation under normal and diseased conditions using organoid cultures. However, heart organoids may still be valuable in studying other aspects of heart chamber development, such as heart pumping and looping.
4. Compact and Trabecular Myocardium Growth
Heart chamber growth was thought to balloon out from the looped hearts segmentally. The ventricular and atrial chambers were found to respectively expand from the linear heart tube on ventral and dorsal sides [
29,
30] and the expanded chambers to develop into two types of myocardium, with the compact myocardium on the outer surface and trabecular myocardium close to the lumen to increase cardiac output and oxygen uptake at early embryonic stages [
31,
32]. The CMs in compact myocardium highly express
Loxl2,
Hey2,
Mycn, and
Fstl4, while the CMs in trabecular myocardium express
Nppa,
Itga6,
Sema3a, and
Slit2 [
32]. Compact and trabecular myocardium development was shown to be differentially regulated by signals from the epicardium and the endocardium [
33,
34,
35]. Epicardium-derived signals such as BMP4, FGF, WNT, IGF, and RA were reported to promote CM proliferation in compact myocardium [
35,
36,
37], and endocardium signals such as NOTCH, Neuregulin, Ephrin, and TGF-β were reported to promote trabecular myocardium development [
31,
38,
39]. Furthermore, some signaling molecules expressed in the myocardium, such as BMP10, were also found to regulate the trabecular myocardium development [
40] (
Figure 1A).
To generate compact CMs from hiPSCs, WNT and IGF2 were added to the ventricular CM differentiation system on day 10 [
41]. The CMs were shown to express typical compact myocardium marker genes such as
HEY2,
MYCN,
TBX10, and
FZD1. In contrast, the addition of Neuregulin to the differentiation system at day 10 can specify CMs into trabecular CMs expressing trabecular myocardium genes such as
NPPA,
NPPB,
BMP10,
IRX3, and
HAS2 (
Figure 1B). Similarly, the co-culture of EndoEC with CMs can promote the development of trabecular CMs, as EndoECs were known to be able to secrete Neuregulin in mice and zebrafish [
42]. Next, it will be interesting to test if other EndoEC-derived growth factors such as TGF-β and NOTCH can also induce trabecular CM fate and if epicardium-derived factors such as RA, BMP, WNT, and FGF can promote compact CM development. Additionally, and most importantly, a test will be needed to determine if these factors can be applied locally to generate heart organoids with compact and trabecular myocardium at correct anatomical locations.
5. Heart Structure Development
The early stages of mouse heart development consist of the formation of the four chambers (left and right atrial; left and right ventricular) and two non-chamber structures—the atrial ventricular canal (AVC) and the outflow tract (OFT). While AVC at later developmental stages contributes to the development of the septum and atrioventricular valves, including the tricuspid and mitral valves, the OFT contributes to the formation of large vessels (aorta and pulmonary artery) and the semilunar valves, including the aortic and pulmonary valves [
43]. The atrial CMs highly express
Nr2f1,
Nr2f2,
Sln, and
Myl7, while the ventricular CMs express
Myl2 and
Mpped2. Furthermore, while the left and right ventricular CMs differentially express
Pcsk6, the left and right atrial CMs highly express
Pitx2 and
Shox2, respectively. In contrast, the AVC CMs express
Rspo3,
Tbx3, and
Bmp2, and the OFT CMs express
Rspo3 and
Cxcl12 [
32,
44] (
Figure 1A).
Atrial lineage specification is regulated by RA signaling in multiple species [
45,
46,
47], while early dorsal-ventral patterning signals such as FGF and BMP also differentially promote atrial and ventricular lineage development in zebrafish [
48]. As the left and right ventricular CMs develop from different heart fields, their lineage formation is primarily regulated by the heart field specification signals previously mentioned when discussing heart fields’ formation. The AVC and OFT share a structure named the endocardial cushion, which is induced by the interaction of BMP signaling, including BMP2 and BMP4 in myocardium and BMPR1A in EndoECs. Mouse endocardial cushion cells express marker genes such as
Twist1,
Msx1, and
Snail. Endocardial cushion cells need to go through an endothelial-to-mesenchymal transition (EndoMT) process regulated by multiple signaling pathways, such as TGFβ, WNT/β-catenin, HIPPO, and NOTCH, to develop into valve cells [
49].
Atrial and Ventricular CMs were found to co-exist in heart organoids but did not display in vivo-like spatial domains. RA signaling had been used to promote atrial CM lineage in monolayer and EB-based hiPSC differentiation, and the atrial and ventricular CM progenitors were distinguished based on the expression of
CD235A and
RALDH2 [
50,
51]. We have also generated heart organoids with atrial or ventricular identities by adding (+) or omitting (−) RA at the cardiac mesoderm stage and found that the RA+ and RA- heart organoids had distinct membrane action potentials and Ca
2+ transient activities. The chamber identity of these cells was further confirmed with immunofluorescence staining for chamber-specific marker genes such as
MYH7,
HEY2 (ventricular),
NR2F2,
MYH6, and
ID2 (atrial) (
Figure 1B). We also performed single-cell mRNA sequencing (scRNA-seq) and random forest-based zone classification to analyze their cell identities systematically [
28]. These analyses consistently support that the CMs in RA- heart organoids preferentially develop into ventricular CMs, while the CMs in RA+ organoids are more likely to develop into atrial CMs [
28].
Interestingly, our study found that
MYL2 is a robust ventricular CM marker gene in human fetal hearts but is barely expressed in the organoid ventricular CMs differentiated from the hiPSC line “WTC”. Considering that WTC and its derived transgenic lines have been broadly used in the cardiac stem cell field, we have investigated
MYL2 expression in WTC-derived CMs from multiple labs based on their scRNA-seq results. We found that while
MYL2 was barely detected in the WTC-derived CMs before differentiation day 30 in several studies, it was expressed in other hiPSC line-derived CMs with the same differentiation conditions [
52,
53,
54]. However, there are some exceptions where
MYL2 was found to be expressed in the WTC-derived CMs on day 30 in one study and day 90 in another. Both studies generated the CMs using a monolayer with small molecules protocol [
53,
55]. The cause of the differences in gene expression across the studies is still a mystery to us, but it will be important to investigate whether this expression variation also exists in other genes and other cell lines.