1. Introduction
The term “autophagy”, (from the Greek words
auto, meaning “self” and
phagein, meaning “to eat”)—literally, eating one’s self—was first created by Christian de Duve over 40 years ago, who discovered lysosomes and provided clear proof of their participation in this process [
1]. It is an evolutionarily conserved process of degradation and recycling in eukaryotic organisms. Two common forms of autophagy have been described in mammals and plants: micro-autophagy and macro-autophagy, while they differ in a third type of autophagy described, chaperone-mediated autophagy (in mammals) and mega-autophagy (in plants) [
2,
3,
4,
5,
6]. The differences among them have been previously described in detail elsewhere [
7,
8], and this review will focus on macro-autophagy (hereafter, autophagy). In this latest process, the cytoplasm and/or organelles are isolated in double membrane vesicles—named autophagosomes—and then transported to the lytic organelle (vacuole in plants and yeast, and lysosome in animals) to be degraded, resulting in the turnover of cellular components. Therefore, autophagy is a fundamental cell clearance pathway that eliminates cellular components, including nucleic acids, proteins, lipids, and organelles, to promote homeostasis, differentiation, development and cell survival.
Autophagy is a unique membrane trafficking process that involves the de novo formation of a membrane, which is generally derived from the endoplasmic reticulum (ER) by generating a double membrane structure called phagophore that elongates to sequester cytoplasmic cargo and closes to form the autophagosome [
6,
9].
The molecular process of autophagy was mostly unknown until 1993, when Yoshinori Oshumi described a genetic screen in yeast, leading to the discovery of AuTophagy-related Genes (ATG) [
10]. 41 yeast ATG genes have been described, and many of them have orthologues in other organisms such as humans and plants.
The autophagy core process in mammals is induced in response to stress by inhibiting the mammalian kinase target of rapamycin (mTOR) or activating 5’ AMP-activated protein kinase (AMPK). In mammals, different stress stimuli can trigger autophagy, such as protein misfolding, hypoxia, nutritional and energy deficiency, ER stress, redox stress, mitochondrial damage, and pathogen infection [
11]. Dysregulated autophagy plays an important role in many pathological processes, including ischemia-reperfusion injury, inflammatory and infectious diseases, obesity and type 2 diabetes, cancer and neurodegenerative diseases [
12,
13,
14].
In plants, inhibition of TOR, usually induced by starvation of nutrients such as nitrogen starvation, is the main pathway that triggers autophagy. In addition, it can also be regulated by repression of glucose signaling, activating the energy sensor Snf1-related protein kinase 1 (SnRK1), which in turn inhibits TOR and activates the ATG1 autophagy initiation complex. The function of AMPK in plant autophagy remains largely unknown, although a plant ortholog of mammalian AMPK, named KIN10, was described as a positive regulator of plant autophagy [
15]. In plant cells, autophagy is triggered by different biotic and abiotic stresses such as oxidative stress, salinity, hypoxia, heat and cold, nutrient starvation, ER stress and pathogen infection. Therefore, autophagy is essential for plants during reproductive and vegetative development, senescence, starvation, immune response and it is critical to cope with environmental stress [
3,
16,
17]. Thus, autophagy must be tightly regulated to maintain cellular homeostasis.
Hydrogen sulfide (H2S) is a colorless, flammable and highly toxic gas known for its rotten egg scent at low concentrations. It has always been considered a toxic pollutant that is found naturally in sewers, stagnant or well waters, compost pits, gas wells and volcanoes. However, it is also endogenously produced in cells by different enzymes.
2. Regulation of Autophagy by Persulfidation in Plants
The role of persulfidation as the molecular mechanism of sulfide for autophagy regulation was first proposed in plants by Gotor’s group [
83] and then several autophagy-related core proteins were demonstrated to be targets for persulfidation [
82,
84]. Recently, the autophagy-related (ATG) proteins, ATG18a, ATG3, ATG5 and ATG7 were published to be targets for persulfidation identified by a quantitative proteomic study in
Arabidopsis leaves [
82]. Nevertheless, the role of persulfidation in those proteins was not deciphered.
In a very recent quantitative proteomic approach in Arabidopsis under nitrogen deprivation, more than 5200 proteins were identified as targets for persulfidation. In this work, authors extended the number of persulfidated proteins involved in autophagy. They found 17 proteins that play an essential role in core autophagy machinery were persulfidated; including ATG2, 3, 4, 5, 7, 11, 13, the serine/threonine kinase TARGET OF RAPAMYCIN (TOR), its effectors proteins REGULATORY-ASSOCIATED PROTEIN OF TOR 1 (RAPTOR 1) and LETHAL WITH SEC THIRTEEN PROTEIN 8 (LST8), five subunits of PP2A, the regulatory subunit of PP2A (TAP46) and the serine/threonine-protein kinase VPS15 [
84]. In addition, this study also revealed that other 58 proteins related to endocytosis and the formation of the phagophore were persulfidated, including several transporters and vacuolar sorting proteins.
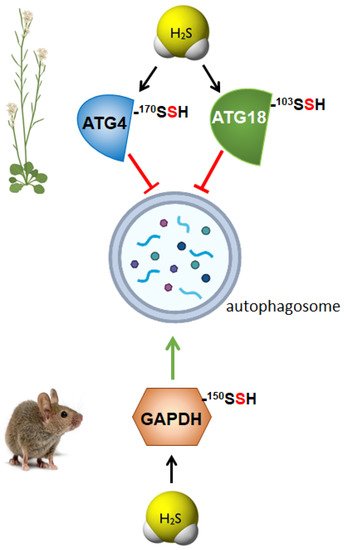
The role of persulfidation as the underlying mechanism regulating autophagy through sulfide was demonstrated in the ATG4 protease, which was specifically modified by persulfidation of Cys170 residue, negatively regulating [
46] (
Figure 2). These authors established that persulfidation of ATG4 upon sulfide treatment inhibited its protease activity, disabling the progress of autophagy. They also revealed that an increase in the intracellular level of the plant hormone abscisic acid (ABA) triggered a decrease in the persulfidated ATG4 level; consequently, its protease activity was enhanced, activating the processing of ATG8, which was further lipidated, and, as a result, autophagy was induced.
Figure 2. Persulfidation of specific proteins differently regulates autophagy in plants and mammals. ATG4, autophagy-related gene 4; ATG18, autophagy-related gene 18; GAPDH, glyceraldehyde-3-phosphate dehydrogenase.
In plants, the role of H
2S in the regulation of autophagy has been studied under stress conditions, particularly under nutrient limitation, demonstrating the negative regulation of bulk autophagy by sulfide through persulfidation of specific targets [
41,
46,
85]. However, a recent research was published describing the role of persulfidation of ATG18a regulating autophagy under ER-stress [
45], and, therefore, deciphering a new level of regulation of selective autophagy through the persulfidation of ATG18a [
45] (
Figure 2). ATG18a binds to phosphoinositides [
86] and forms a complex with ATG2, which is involved in autophagosome biogenesis during phagophore expansion [
87]. In this research, it was demonstrated that sulfide regulates ATG18a phospholipid-binding activity by reversible persulfidation at the specific residue Cys103, which reversibly activates ATG18a binding capacity to specific phospholipids. Authors proved that the mutation of Cys103 in ATG18a decreased its binding capacity to membranes and its localization time within the phagophore was shorter. In this way, the reversible persulfidation of ATG18a affects its binding to membranes, which potentially delays its release from the autophagosome, inhibiting autophagosome progression and maturation [
45]. This regulation of autophagy through the persulfidation of ATG18a probably allows the plant a correct physiological response upon stress, with the final outcome of plant survival.
3. Regulation of Autophagy by Persulfidation in Animals
The molecular mechanism by which H
2S regulates autophagy in mammals has been recently established thought the study of the persulfidation of glycolytic glyceraldehyde-3-phosphate dehydrogenase (GAPDH) [
88] (
Figure 2). This modification of GAPDH triggers its translocation to the nucleus, which is critical to induce autophagy via deacetylation of the autophagic core protein LC3B, and the consequential autophagosome formation. Authors demonstrated that nuclear GAPDH interacts with the cell cycle activator and apoptosis regulator 2 (CCAR2/DBC1), avoiding the interaction of CCAR2 with deacetylase SIRT1, and therefore avoiding the inactivation of SIRT1. Then, activated SIRT1 deacetylates MAP1LC3B/LC3B (microtubule-associated protein 1 light chain 3 beta) inducing its translocation into the cytoplasm and activating autophagy [
88]. Persulfidation of GAPDH at the same residue, Cys150 was previously described in mammals resulting in an increase of its enzymatic activity [
76], although in later studies, polysulfide treatment of GAPDH showed opposite effects decreasing its enzymatic activity [
89]. In plants, persulfidation of the cytosolic isoform GAPDH (GapC) was also previously described [
78] and demonstrated its nuclear translocation upon the modification of the protein [
77]. However, in none of these studies, the relationship between persulfidation of GAPDH and autophagy regulation was analyzed. A similar situation comes about the regulation of protein tyrosine phosphatase (PTP1B), which was described to be persulfidated at Cys215, inhibiting its enzymatic activity. This inhibition resulted in the activation of PERK alleviating ER stress [
66], but authors did not relate this regulation to autophagy. Nevertheless, later was demonstrated that activation of PERK by sulfide treatments, increased eIF2 phosphorylation and induced autophagy [
68], probably due to the persulfidation of PTP1B.